Vegetation of the Ruahine Range

THE FOREST RESEARCH INSTITUTE
THE functions of the Forest Research Institute are to supervise all forestry and forest-products research carried out by the New Zealand Forest Service, and to undertake a programme of research approved by the Director-General of Forests. The work of the Institute is organised under six branches: Silviculture, Pathology, Forest Management, Protection Forestry, Forest-tree Improvement, and Forest Products. Most of the professional staff, together with field and auxiliary-service personnel, are based on Rotorua, but subsidiary Experiment Stations are located at Rangiora, Napier, and Wellington. All come under the charge of the Director of Research, who is directly responsible to the Director-General of Forests.
The work of the Institute is reported in its Annual Report and through the following media: New Zealand Forestry Research Notes, New Zealand Forest Service Technical Papers, and New Zealand Forest Service Bulletins. Papers published in scientific journals may be issued in a Reprint series. These publications are available from either the Director-General of Forests, Private Bag, Wellington, or the Director of Research, Private Bag, Whakarewarewa, Rotorua.
S.D. RICHARDSON,
Director of Research

TRANSACTIONS
OF THE
ROYAL SOCIETY OF NEW ZEALAND
BOTANY
VOL. 3 No. 3 APRIL 14, 1965
Vegetation of the Ruahine Range: An Introduction
By N.L. ELDER
(Received by the Editor, 25 March 1964.)
Abstract
THE Ruahine Range forms a vegetational bridge between the volcanic plateau and Cook Strait, that is from the driest and sunniest environment in the North Island to one of the cloudiest and wettest. Past and present changes are conspicuous and their causes various. This paper summarizes observations over the last 30 years and is presented in the form of an introduction to the more specialized and detailed surveys now being carried out by the New Zealand Forest Service.
LOCALITY
THE Ruahine Range is that portion of the main North Island fold mountains which lies between the Manawatu and Taruarau-Ngaruroro gorge systems. These separate it from the Tararua Range to the south and the Kaweka Range to the north. (Map 1.)
The Ruahine Range extends 56 miles in a NNE direction from the Manawatu Gorge, and is comparatively narrow, the southern third having an average width of little more than five miles, then widening to a maximum of some 15 miles towards the northern end. There are few foothills, and on the eastern side the divide forms a high continuous scarp above the plains of Hawke’s Bay, but there are a number of high subsidiary ranges diverging from the divide on the western side.
On either side farmlands extend to the foot of the main slopes, and at the southern end farmlands separate the range from the Manawatu Gorge itself. The induced scrubland of the Taruarau Gap intervenes between the Ruahine and Kaweka Ranges.
STRUCTURE
It is thus a high (5,678ft max.) continuous narrow range cut off from the rest of the mountain system. Its eastern boundary is formed by the Hawke’s Bay fault system, its western by the echeloned valleys of the Rangitikei, Oroua, and Pohangina Rivers and its northern and southern boundaries by the traverse gorges already mentioned.
Published by the Royal Society of New Zealand, c/o Victoria University of Wellington, P.O. Box 196, Wellington.

14 Transactions – BOTANY Vol. 3.
In profile as seen from the eastern side the range rises in notably smooth and even slopes from the gorges at either extremity up to an altitude of 4,000 – 4,500ft above sea level. The outline then becomes broken, but its peaks continue this gradient until the central block of the range is reached when there is a well marked step up of 500 feet or so. All points approaching or exceeding 5,500ft lie in this central block.
From the western side the profile does not show such distinct features. From the central block several high subsidiary ranges diverge in a NNW direction and hide most of the main divide. South of this divide between the Pohangina and Manawatu Rivers swings east and falls away to a scrubby ridge which is completely hidden from the west by two echeloned side ranges with a southerly trend. South of these again the divide rises to the oblong block of the Southern Ruahine Range.
The structure of the northern end of the Ruahine Range is more complicated. The greywacke axis terminates in a plateau overlooking the Taruarau Gorge and is about the only coherent geological features here in a crazy pavement of faulted blocks; the actual divide lies across the Pohokura basin from this, at first following the limestone scarp of the Mangaohane plateau, then continuing along a greywacke outlier, the OtupaeRange, to abut on the Ngamatea plateau which separates the Rangitikei and Ngaruroro catchments.
As described by Kingma (1959) the range has been elevated in recent geological times with Tertiary limestones occurring intermittently across its northern end and capping spurs in the Makaroro valley. It is probably moving as a series of transcurrent faults southward relatively to the Kaweka and Kaimanawa Ranges, leaving the Taruarau Gap and on a lesser scale the Pohokura basin and Colenso’s Lake as collapsed blocks, the latter two of especially stratigraphic interest as preserving the sequence of the original Tertiary deposits.
SURFACE WINDS 0900 NZST 1940-1949 (percentages)
calm 10% TAIHAPE
calm 10% HASTINGS
Wind directions
TEXT-FIG. 1. – Wind Polygons (9.00 a.m.) showing contrasting wind patterns on opposite sides of the range.

No. 3 ELDER – Vegetation of the Ruahine Range 15
Although steep slopes and extensive screes are characteristic of the eastern face of the range, where they may be assumed to be mainly tectonic in origin (Plate 1), as well as several of the inner valleys of the central block, wide rolling crests and plateaux are characteristic particularly on the western wide. The sculpture of the Ruahine comes between that of the Kaimanawa and that of the Tararua, being further advanced than the former but not carried as far as the latter.
VOLCANIC ASH
Deposits of volcanic ash are negligible in amount except for remnant pumice hummocks of up to 3ft in depth on the lower part of the NE plateau, apparently an accumulation of wind drift. Two deposits can be distinguished in peat on the NE and Mokai Patea plateaus and as far south as Whanahuia. The most recent shower here is 3in thick, but beyond this both decrease sharply in thickness. Below the Waimihia ash, to the full depth of the peat on the No Man’s bog, are six or seven narrow separate ash bands, but there is no indication of a massive “Tongariro” shower.
CLIMATE
In general terms over 40% of the winds in the central areas of New Zealand have a westerly component, and over 20% come from between south and east; but these directions are considerably modified by topography, so that the differences on either side of the Ruahine Range are considerable. (Text-fig. 1.)
Nor’westerlies and westerlies in the South Taranaki Bight converge into the wind funnel of Cook Strait, with a change of direction tending to follow the coastline. Inland of this, towards the barrier of the Ruahine Range westerly-wind velocities appear to drop, and only occasionally do these winds cross the range with the development of a characteristic Fohn arch. The Manawatu Gorge, however, forms a minor wind funnel for westerly winds, bringing a high proportion of cloud to the lower slopes north and south of it.
On the Hawke’s Bay side of the range sou’westerlies and southerlies supply most of the cloud and most of the rain days, though the heaviest falls are brought by the comparatively infrequent winds between NE and SE.
The Taruarau Gap appears to have no effect on wind patterns, unless perhaps it contributes to the anomalous NE component of the Taihape wind polygon.
RAIN. Except for some gauges that the Hawke’s Bay Catchment Board have recently installed on or near the divide, rainfall is measured on the margins of the range only. The distribution of rainfall in the Ruahine Range is therefore at present not known.
Figures are low at the northern end, Mangaohane averaging below 40in though it is at an altitude of 2,750ft, and Whanawhana, opposite it at the eastern end of the Taruarau gap, just over 40in. (North of this, however, at Kuripapango, the rainfall approaches 60in in the lee of the Kaweka Range.) A slight increase appears to occur towards the south, reaching 50in at Rangiwahia on one side of the range and the same at Blackburn on the other. There is then a considerable increase, Table Flat, where the Oroua leaves the range, having 75in, and a new station (1,670ft) behind Ashley Clinton, probably over 90in, while south of this again Umutaoroa has 100in. No information is yet available from a gauge established on the divide (4,353ft) at the head of the Waipawa River.

16 Transactions – BOTANY VOL. 3
SNOW. The heaviest snowfalls appear to visit the western side of the range. Colenso mentions snow damage on one of his early journeys, but damage to forest from this cause is not typical. The series of heavy snowfalls of the winter of 1961 were quite exceptional. They lay for a period and one particularly heavy fall, probably early in August, did considerable damage, particularly in red beech forest.
CLOUD. Cloud levels have been observed (9 a.m., 4 p.m.) from Havelock North along most of the eastern side of the range over a period of three years, July 1938 to August 1941, and over 13 months of this period observations from Kumeroa of the southern part of the range were also taken, together with a short check period of a month from Dannevirke.
The preponderance in Hawke’s Bay of winds with a southerly component has an effect on the cloud cover in the northern half of the range. From observations made from Havelock North the peaks at the head of the Oroua River, where the highest part of the divide begins, 25% of the days were reckoned to be substantially sunny. On the same basis this increased to 30% at the northern end of the range, while further north again in the Kaweka Range it reached 34%, the greatest value observed.
The southern half of the range is lower and the weather is increasingly dominated by a westerly component as the gap of the Manawatu Gorge is approached, with a considerable decrease in sunny days and increase of low cloud, and with these a marked change in the vegetation. The transition is difficult to observe from the Hawke’s Bay Plains, as the higher Ngamoko Range to the west of the divide shields it from the westerly winds but is largely concealed by it. South of the Pohangina Gorge, however, low cloud is characteristic of the Southern Ruahine, averaging 67% on Maharahara at about its centre and even higher on Wharite, the southernmost peak, both averaging only some 11% of sunny days over a 13-month period.
No comparable information is available for the western side of the range, but from sporadic observation cloud or fog frequently forms or lies along the western slopes when the rest of the range is clear. The different composition of the higher forest on these outer faces suggests that diminished sunshine is a factor.
TEMPERATURE. Both summer (63° F.) and winter (46° F.) isotherms run N-S along the general line of the range, the Hawke’s Bay side being warmer in summer and cooler in winter than the western side.
HISTORY
The Ruahines, like other ranges, were visited by the Maoris when hunting for birds (the skeleton of a lost hunter was noted by Colenso near Puketaramea) and as refugees (several traditional sites are recorded in the headwaters of the Mangatera); a number of traditional high-country routes are on record. As the Ruahines are a comparatively long narrow range, cross-country routes to and from Hawke’s Bay and the Inland Patea were of particular importance (the Te Atua Mahuru route, Te Parapara, being used by a war party as late as 1828), and there were also routes, less certainly known today, between southern Hawke’s Bay and the lower Rangitikei. In addition traditional routes ran north and south along the open crests in the northern part of the range. The more exposed routes could obviously be used only in favourable weather for lack of suitable clothing, and groups of tarns, which are a certain source of water even in periods of prolonged dry weather,

No. 3 ELDER – Vegetation of the Ruahine Range 17
have traditional names. It has been suggested that muttonbirds, which were formerly known to nest above the bush line in considerable numbers, would have afforded a seasonal food supply of considerable value to parties making high-level traverses. The main effect on the vegetation of the use of the high country by Maoris lies in their use of fire for hunting or track marking, and several extensive areas of old fires suggest such fires that had got out of control. However, by no means all Maori routes took advantage of the open snowgrass tops; the best known (Te Parapara) followed valleys on either side, reducing travel above the bush line to a minimum. Colenso records the tradition of the destruction of a war-party here on a snow slide coming off Te Atua Mahuru.
This route is of particular botanical interest, as Colenso ascended it in 1845, making the first collection for the whole of New Zealand of a large number of high-country species. The listing of type localities in Allan’s Flora Vol. I (1961) gives at least 80 species and accepted varieties, exclusive of monocotyledons for the Ruahine Range and its neighbourhood.
Since Colenso’s time a certain amount of sporadic botanical collection appears to have been done along the flanks of the range, but it is not until after 1900 that there is any further record of extensive journeys, the most important being that of F. Hutchinson and B. C. Aston retracing Colenso’s route in 1911.
A more methodical botanical exploration of the range began in 1933 and, as a result of this, comprehensive vegetation maps were drafted by Druce in 1946, summarizing the coverage that had been made up to the break caused by the war. In the last 10 years various surveys have been carried out by different branches of the Forest Service with the aid of aerial photographs. This paper is for the most part a summary of the earlier work and essentially an introduction to those more detailed surveys.
FAUNA
BIRDS. There is a rumour or tradition that huias survived in the NW Ruahine up to about 1907, and a story is on record (Phillipps) of a skin collected in the same area by Morgan Carkeek in 1894 which was recognised as that of a “mohoau” (Notornis). Wekas were last seen at Kereru about 1925 (L. Masters, pers. comm.).
The narrowness of the range makes it a poor refuge, and visitors over at least the past 25 years have been repeatedly struck by the apparent paucity of bird life. The restriction of seasonal movement with the general replacement of flanking podocarp-hardwood forest by grassland is probably a sufficient cause. The winter migration of tuis to the eucalypts of the Heretaunga plains has been regular for at least the last 30 years, and recently bellbirds and, more surprisingly, pigeons and kakas have wintered in Havelock North, 30 miles or more from the nearest extensive forest. Some introduced birds are widely distributed throughout the range, the blackbird and the chaffinch being the most conspicuous; others are mainly marginal, the Australian magpie, for example, being strictly confined to grassland. Chaffinches have been noted flocking on occasion, but otherwise the paucity of bird life extends to introduced species.
This apparent scarcity of bird life needs confirmation. Some assessments are being made in the course of forest surveys (Ellis). If true it must be expected to have a bearing on insect control and on seed distribution and/or consumption. The only evidence of this nature comes from the Pohangina valley, where local

18 Transactions – BOTANY VOL. 3
residents reported heavy defoliation of kowhai along the river flats by pigeons. The most likely reason for this is that competition from opossums in the adjacent Southern Ruahine had forced the pigeons on to alternative winter browse. The heavy flowering of kowhai in the Pohangina since that report suggests that the pressure has since eased, the opossum population being lower since the peak and collapse of mid 1955.
MAMMALS. The comparatively accessible tussock country of the northern and western Ruahines was stocked in the 1880’s, in the first place with merino sheep, and this was accompanied by more or less intense tussock burning. The NE plateau and the Otupae Range have been abandoned, apart from strays, for 20 years or more, but stock, mainly cattle, are still run on Mangaohane and the Mokai Patea, while the Pohokura basin has recently (1956) been disced and sown down in grass.
Towards the southern end of the range the practice of wintering cattle in the bush has not entirely died out, and names such as “The Cow Saddle” recall a former Te Ohu Station mustering route across the divide.
When the present survey began in 1932 the effect of red deer was inconspicuous except in the NE comer of the range, while opossums and goats were practically absent. As the survey proceeded, information about the animals, their spread and, later, measures for their control, together with observations on their effects on the vegetation became an integral part of what had begun as a botanical survey. It forms, however, an unwieldy component of the paper in its present form and for neatness’ sake has been treated separately (Appendix 1).
FIRE
There is some evidence of pre-European fires at three points in the north and west of the range. On the Mokai Patea plateau the name Tahu-a-Rongotea (now given in the shortened form, Rongotea, to its culminating point) refers to fire. Early maps show that it was given to an area about half way down the plateau; and the irregular and unusual timber line on the eastern slope, together with, across the valley, an anomalous scrub face bearing a Maori name, Otukota, is supporting evidence of fire (Plate 2). Unfortunately there is no legendary information from which an approximate date might be estimated, but the recent discovery of buried charcoal, resembling deposits that are being studied in Canterbury, indicates that it occurred in the early stages of Maori occupation of the country (Wardle, pers. comm.).
A fire pattern extending from the margin of the Mangaohane tussock down the Otorere tributary of the Mangatera could well be thought, from a distant view, to be a consequence of European burning; but from the size of the regeneration dominants (Phyllocladus) this must be reckoned to be of considerable age.
The Pohokura basin may be considered a third area affected by pre-European fire as it is first recorded as occupied by tall kanuka (Leptospermum ericoides) scrub, a highly unlikely climax in a low-lying sheltered limestone basin with occasional pockets of matai (Podocarpus spicatus) forest.
The Inland Patea tussock was occupied at an early stage of European settlement, and a good deal of the isolated Otupae Range carries young beech (Nothofagus) forest which has partly regenerated after burning of that date. Although evidence of tussock burning is general it should be put on record that at one period Mangaohane policy was against burning (Morrin).

No. 3 ELDER – Vegetation of the Ruahine Range 19
On the eastern side of the range the original Seventy-mile Bush, the belt of podocarp-dominant forest that ran between the foot of the range and the tussock-covered Takapau and Ruataniwha Plains from the head of the .Manawatu River to Kereru has been almost entirely replaced by grassland, its outline being roughly preserved by the scattered totara (Podocarpus totara, see Appendix 2) of bushy form which appears to have been able to regenerate m grassland m the face of browsing.
Fires have here and there run up the face of the range, and in some instances have been more extensive. In 1910 a fire ran up the Makaroro valley for several miles as far as the start of Colenso’s Track, and in 1939 fire in cut-over bush adjacent to this in the Makaroro mill workings has left an area of several hundred acres partly in grassland, partly in dense wineberry (Aristotelia serrata) thickets which are still persisting.
There are traces of fire on both sides of the northern head of the Waipawa River, and if even-aged regeneration of mountain beech (Nothofagus solandri var. cliffortioides) on old screes in the basin is connected with this, as seems likely, an extensive fire occurred here prior to 1903.
Two major fires have occurred near the head of the Tukituki, the first, some time in the 1880’s, spreading from either Howlett’s Hut or the head of the Moorcock Stream, across the range into the head of the Oroua and further south across the Pohangina Saddle and into the head of the Makeretu River, an area of some 5,000 to 6,000 acres. The name Daphne Ridge is said to come from the abundance of Pimelea buxifolia in the scrub-tussock succession that followed the burn
During the drought of 1946 a fire starting in the Pohangina Hut ran over part of this area, reaching the divide just south of Otumore, but mainly travelling back into the head of the Moorcock valley.
In the Southern Ruahine there are traces of fire on the lower forest margins and up some of the outer spurs, characteristically indicated by dense stands of Cyathea medullaris, the large black tree fem, but none of these is extensive. One fire, however, in 1915 ran up the Pohangina Gorge from the west, crossed the range at the Apiti-Norsewood saddle, and went on to Te Ohu Station. My information comes from L. H. Bowden, who stresses that this was a series of spot fires and that the bush in between was untouched. Today a few anomalous patches of Coprosma/Myrsine scrub in the forest near this saddle presumably mark the sites of these spot fires.
An extensive fire which is stated to have run southwards along the western margin of the Southern Ruahine about 1927 appears to have been confined to farm land and to have had little effect on the main forest.
VEGETATION
TAXONOMY. For so comparatively limited an area the Ruahine Range has become the type locality of an exceptional number of high-country plants, thanks to Colenso’s early missionary visits to the Inland Patea. The regular listing of type specimens and localities in the 1961 Flora is, because of this, of particular assistance to work in the Ruahine Range. In addition, the provision of footnotes explaining the history of the nomenclature and the status of the various synonyms and partial synonyms which have bedevilled taxonomists in the past is most welcome, as so many of these originated with Colenso’s collections in his later

20 Transactions – BOTANY VOL. 3
years in the remnants of the Forty-mile Bush around Dannevirke. Some of these can usefully be distinguished as true-breeding forms when one is working on a local scale. Names that are in popular use for certain widespread or characteristic species are used in the text. Their formal equivalents are listed in Appendix 2.
SPORADIC DISTRIBUTION. The Flora records geographical distribution in general terms. Consequently a number of species, such as those occurring on the volcanic plateau and again in NW Nelson, are given a range which includes the Ruahine, though they have not in fact been observed there. Others are known only from one small colony, as that of semi-juvenile Pittosporum turneri (Mangaohane forest margin), or even from solitary plants, as P. colensoi (Oroua) and Phymatodes novae-zelandiae (Pohangina) at considerable distances beyond their previously known distribution. There are probably others, but none of these is likely to prove of any significance in their communities. This sporadic occurrence applies to two species for which the Ruahine Range is the type locality, Lycopodium novae zelandicum and Nothofagus truncata, neither of which has been recorded in field surveys in the past 30 years; in fact there is a gap in the known occurrence of the Nothofagus at the present day of 100 miles or more along the East Coast. Ascarina lucida may be mentioned here. Its pollen has been recognized in a Western Ruahine bog though the living plant has not been recorded.
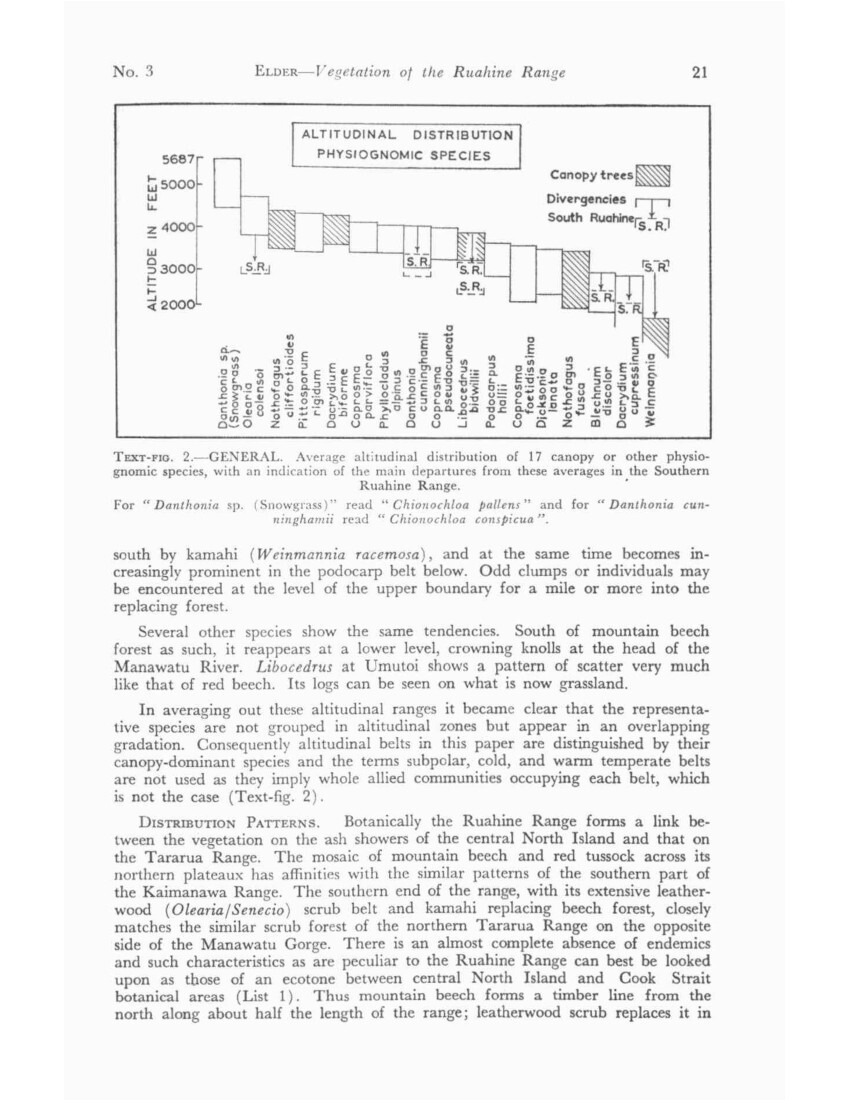
ALTITUDINAL BELTS. The most striking feature of high-country vegetation is its stratification into well marked altitudinal belts, and the examination of these is the most obvious line for investigation of distribution. These belts were used as the basis of an earlier survey of the Tararua Range, but there was a suspicion there that there had been a tendency to lump groups of species into belts on rather too arbitrary a basis. Therefore from the commencement of work in the Ruahine care has been taken to check on this tendency by establishing a network of corrected spot heights and noting the altitude at which different species occur. Frequencies also have been recorded on a scale which, though necessarily approximate, is as consistent as it can be made on a personal estimate. Field experience with Forest Service survey plots has afforded a valuable check on the accuracy, within limitations, of such estimates.
Analysis of these records for a selection of the more important species shows, of course, variations recognizably due to exposure, aspect, and presumably in some instances to soils, but these average out to give mean altitudinal ranges which can be accepted with confidence. Upper limits are very sharp and in practice it is fairly easy to select a line above which any given species ceases to be a recognizable component of its community; occasional individuals which may occur above this are most commonly limited to a vertical interval of the order of as little as 200ft.
The corresponding lower limit is usually less sharply defined and the sporadic scatter of individuals of a species to lower altitudes is wider. However, sufficient information is available to indicate that the vertical range of nearly all the main species examined (that is to say, the range within which they can be classed as a definite component of their community) is with quite striking constancy, between 900 and 1,200ft.
Red beech (Nothofagus fusca), on first calculation, was the chief exception, with some wide downward fluctuations below a fairly regular upper boundary. A re-examination of observations showed that these were associated with the absence of red beech from both ends of the range. In either case as it approaches its limit it is increasingly replaced, in the north by mountain beech and in the

No. 3 ELDER – Vegetation of the Ruahine Range 21
south by kamahi (Weinmannia racemosa), and at the same time becomes increasingly prominent in the podocarp belt below. Odd clumps or individuals may be encountered at the level of the upper boundary for a mile or more into the replacing forest.
Several other species show the same tendencies. South of mountain beech forest as such, it reappears at a lower level, crowning knolls at the head of the Manawatu River. Libocedrus at Umutoi shows a pattern of scatter very much like that of red beech. Its logs can be seen on what is now grassland.
In averaging out these altitudinal ranges it became clear that the representative species are not grouped in altitudinal zones but appear in an overlapping gradation. Consequently altitudinal belts in this paper are distinguished by their canopy-dominant species and the terms subpolar, cold, and warm temperate belts are not used as they imply whole allied communities occupying each belt, which is not the case (Text-fig. 2).
DISTRIBUTION PATTERNS. Botanically the Ruahine Range forms a link between the vegetation on the ash showers of the central North Island and that on the Tararua Range. The mosaic of mountain beech and red tussock across its northern plateaux has affinities with the similar patterns of the southern part of the Kaimanawa Range. The southern end of the range, with its extensive leatherwood (Olearia/Senecio) scrub belt and kamahi replacing beech forest, closely matches the similar scrub forest of the northern Tararua Range on the opposite side of the Manawatu Gorge. There is an almost complete absence of endemics and such characteristics as are peculiar to the Ruahine Range can best be looked upon as those of an ecotone between central North Island and Cook Strait botanical areas (List 1). Thus mountain beech forms a timber line from the north along about half the length of the range; leatherwood scrub replaces it in
Photo caption –
ALTITUDINAL DISTRIBUTIONPHYSIOGNOMIC SPECIES
TEXT-FIG. 2. – GENERAL. Average altitudinal distribution of 17 canopy or other physiognomic species, with an indication of the main departures from these average in the Southern Ruahine Range.
For “Danthonia sp. (Snowgrass): read “Chionochloa pallens” and for “Danthonia cunninghamii read “Chionochloa conspicua”.
22 Transactions – BOTANY VOL. 3
the south and it overlaps it in the centre but is absent from the northern quarter of the range. Both are physiognomically important and with associated species enable natural areas to be distinguished in a north-south direction along the main backbone of the range.
There is also an east-west ecotone running from this main axis across the subsidiary ranges which run out on the western side. Mountain beech does not reach the western margin; its place is taken by cedar (Libocedrus) forest, an extension of the cedar forest which occurs in a broken pattern from the margin of the Waimarino Plain, round the western side of the volcanic plateau and across the south-western extremity of the Kaimanawa Ranges.
Thus the timber line, with or without a scrub line, shows several distinct patterns both from north to south and from east to west, and the presence or absence of red beech and kamahi at lower altitudes also shows patterns which correspond to some of these and together form a basis for a subdivision of the range into natural areas.
NATURAL AREAS (Map 2)
These may be listed in their simplest form as follows:
CENTRAL RUAHINE. Mountain beech dominant above 3,600ft, red beech below this.
NORTHERN RUAHINE. Mountain beech dominant or alternating with red tussock, red beech scarce.
WESTERN RUAHINE Cedar dominant above 3,600ft, red beech below.
SOUTHERN RUAHINE. Leatherwood dominant above 3,000ft, kamahi below this. Beeches absent.
The ecotone between Central and Southern Ruahine forms a belt about six miles wide which runs diagonally across the Ngamoko Range, and, though here included in the Western Ruahine, may be considered as a further subdivision. Though mountain beech is absent, it differs from the Western Ruahine in so far as Dacrydium biforme tends to replace cedar at the timber line and dominates it on the eastern side.
The Central Ruahine is considered first as the basic pattern of which the surrounding areas are variants.
CENTRAL RUAHINE (Map 4)
On this basis the Central Ruahine includes all points on the divide exceeding 5,500ft and most of the diverging ranges to the west of it between the Mangatera and Pourangaki valleys, so that about a third of the area lies above the timber line and the only considerable areas of high country outside its boundaries are the Whanahuia and Ngamoko Ranges to the south, carrying a western type of vegetation.
The boundaries are on the whole fairly satisfactory. The northern boundary cuts off the plateau country which terminates at the head of the Makaroro valley; the southern boundary slanting across the Pourangaki and Oroua valleys marks fairly closely the replacement of mountain beech timber line by cedar. The western boundary also follows the change in timber line but is less satisfactory in that the

No. 3 ELDER – Vegetation of the Ruahine Range 23
tussock and bog of the Mokai Patea would more conveniently be grouped with those of the Whanahuia Range, bringing out certain affinities between the western and northern subdivisions.
The eastern face of the range drops abruptly 3,000 to 3,500ft on to the plains of Central Hawke’s Bay, and contains only two longitudinal valleys of any size, the Makaroro and Tukituki, so that forest on this side occupies a fairly narrow belt along the face of the range; west of the divide larger forested areas occupy the Mangatera and Maropea basins. The upper portions of these as well as of the Kawhatau and Pourangaki valleys fall within the area of mountain beech forest.
CENTRAL: PODOCARP FOREST
In the course of European settlement, the whole of the Hawke’s Bay (eastern) side of the range has been cleared and grassed up to the foot of the main scarp, to an altitude, opposite the middle of the range, of about 2,000ft. Consequently only traces remain of lowland forest, the former belt of podocarp-maire (Gymnelaea spp.), the extension of the Forty-mile Bush known as the Seventy-mile Bush, which ran between the tussock of the Takapau and Ruataniwha Plans and the Ruahine Range to as far north as Kereru. From the few remaining pockets, of which Gwavas Bush is the largest and most important survivor, totara was an important constituent in places and the regeneration of totara in grassland which is now a feature of the rolling country from Ashley Clinton to Kereru appears to outline the area what was previously forested, although Podocarpus totara has not been seen within the boundary of the range in the vicinity. (List 2.)
At the present day a fringe of podocarps, mainly rimu (Dacrydium cupressinum) with a little matai (Podocarpus spicatus) and kahikatea (P. dacrydioides) occur either scattered through the lower red-beech forest or in small pockets on favourable sites (List 3A). Scattered trees of miro (P. ferrugineus) are also present to somewhat higher altitudes, while thin-barked totara (Podocarpus hallii) is rather a component of red-beech forest, whose altitudinal range it matches, than of podocarp forest proper. Logging in these remnants at the northern (Makaroro) and southern (Moorcock) margins has only ceased in the past few years.
In the heart of the range at Colenso’s Lake (2,300ft) which lies on a down-faulted tertiary block in the upper Mangatera valley, podocarps occur in red-beech-dominant forest together with a number of characteristically lowland species. The abundance of matai and relative scarcity of rimu relate this community to the matai-dominant podocarp remnants in the Northern Ruahine. Although totara has been reported from Colenso’s Lake this appears likely to have been a misidentification as only the related Podocarpus hallii could be found on a recent visit.
CENTRAL: RIVER BANK
The streams in the larger valleys, particularly those draining the eastern side of the range, are unstable at the present ay with frequent changes of channel and much deposition of shingle and scouring out of older terraces, which suggest at least one previous cycle of activity. An occasional local cloudburst, as that of the winter of 1954 in the Makaroro-Waipawa area, brings about major changes which will be recognizable over a long period.

24 Transactions – BOTANY VOL. 3
A succession of specialized plants (List 4) colonizes the bare shingle, beginning with mat plants, continuing with shrubs and certain trees, the more stable terraces eventually being occupied by the neighbouring forest species. Some species which may not be found in the main forest, such as kowhai (Sophora tetraptera) and kamahi, commonly occur on the hill slopes immediately above the terraces, probably favoured by the increased light that reaches them there. A sprinkling of high-altitude species that have come down stream is characteristic.
River cliffs, both dry and wet, also support characteristic communities, including at the resent day a number of plants that are palatable to browsing animals and which have almost disappeared from accessible sites.
A few species in these categories but otherwise confined to the Northern Ruahine extend into the central area, the Central North Island form of Hebe parviflora var. arborea on river terraces in the Makaroro on cliffs Carmichaelia odorata var. pilosa in one of the Makaroro tributaries, and Angelica rosaefolia in the Makaroro, Maropea, and Mangatera valleys.
CENTRAL: RED BEECH FOREST (List 3)
From 2,000ft upwards red beech is the usual dominant and above the limit of rimu (c. 2,800ft) it is the sole canopy dominant up to 3,600ft, higher in sheltered basins and on deeper soils, lower on exposed ridges. Closely associated with it is Podocarpus hallii with an almost identical altitudinal range, but usually, in this type of forest, as a sub-canopy species. As has been noted in describing podocarp forest, miro overlaps into red-beech forest as a minor component. A fair amount of light filters through the canopy and over wide areas one or other of two species of fern, Dicksonia lanata or Blechnum discolor forms a dense waisthigh floor cover. Few shrubs can compete with this cover and seedlings beneath it are infrequent: Neither fern appears to show any site preference over the other and it is exceptional for either to show any trace of browsing. The Dicksonia, until the new Flora appeared, was listed as of infrequent occurrence- “somewhat scarce and local in the North Island” (Dobbie, 1951). Its abundance at the [the] present day over wide areas in several of the central ranges may be a recent development and the opening of the floor by deer a possible factor.
North-facing slopes on which scattered large red beech form a discontinuous canopy with no replacement in sight (pole, sapling, or seedling) and with a dense fern cover beneath them are frequent in the Ruahine, as in the northern part of the Kaweka Range. As neither fern species flourishes under full exposure to light the further opening of the canopy may be expected to give rise to major changes, but it is not yet clear what these will be.
Towards the northern boundary of the area – that is to say, approaching the belt of country across the Inland Patea where red beech occurs sporadically or is absent from wide areas, it tends to persist at low altitudes but is increasingly replaced by mountain beech above 3,000ft. In several localities where it persists at ca. 3,600ft a canopy of stag-headed red beech with mountain-beech saplings and seedlings beneath indicates a process of replacement.
Towards the southern boundary sub-canopy kamahi is an increasingly important component of red-beech forest, becoming co-dominant beyond this and replacing it in the Southern Ruahine.

No. 3 ELDER – Vegetation of the Ruahine Range 25
Some islands of mountain-beech forest below 3,000ft are conspicuous in aerial photographs, chiefly between the Makaroro and Waipawa Rivers. Their dense canopy suggests even-aged fire patterns, but investigation has shown no trace of fire, and in one of them at least on the Shut Eye Ridge not only is the mountain beech all-aged, up to 125 years old, but several large red beech, up to 40in in diameter, are scattered through with their trunks shorn off below canopy level and their crowns merging with and indistinguishable from it. These islands occur on knobs and ridges, the even canopy is a wind roof and the pattern probably originated from a destructive gale more than 120 years ago.
One feature in red-beech forest on steep valley faces has been the increase in the number of slip faces, first apparent about 1940 in the Waipawa and Makaroro, where their development has been most closely followed. The increase can fairly certainly be attributed to deer, which reached their peak in this area about 1935. Slips frequently started where deer had congregated on sunny slopes, their trampling destroying the moss and root cover, after which the exposed rock began to disintegrate. Recent observation (1962) shows the beginning of revegetation on several of these in the Waipawa, and fresh rock falls only on a minor scale; intensive hunting had then been carried on for three years in this valley and seems to be the major reason for this recovery.
In the Makaroro one large scree blocked the river about 1957 and formed a lake which persisted for two or three years; a similar dam across the Maropea River was encountered in 1941.
In the Makaroro deer damage is only part of the story. The valley sides are steep and present a series of comparatively stable faces or bluffs with unstable rubble faces between them. The fire of 1910 swept these slopes in the lower part of the valley and today second-growth scrub forest with a good deal of exposed rock is common below Colenso’s Track. Upstream of this, however, although red beech is dominant, with exceptionally vigorous regeneration on stable slopes, the rubble slopes are usually occupied by Pittosporum eugenioides, Melicytus ramiflorus, or Aristotelia serrata, the latter sometimes in pure stands. The floor commonly consists of loose boulders or rubble and large beech logs are frequent. Clumps of pole red beech occur sporadically, but otherwise over most of these faces the absence of beech regeneration is marked. The pattern does not suggest fire and no trace of charring has been found in the upper part of the valley.
However, some catastrophic event seems to have been responsible, and the abundant evidence of recent faulting in the Makaroro headwaters, some of which is conspicuous in aerial photographs, suggested that a major earthquake within the last 200-300 years was a likely cause.
The most conspicuous and apparently most recent fault trace which shows up as a narrow furrow in the canopy a mile or more in length along the western slope of the Pohatuhaha Range, was examined with this possibility in mind, but the presence of large cedar, reckoned to be 400-500 years old, growing in the actual fault trench, fails to bear this out (Plate 3).
CENTRAL: MOUNTAIN BEECH FOREST
Above red beech (c. 3,600ft) mountain beech is the dominant canopy species to the timber line, which has an average maximum altitude of 4,400ft, but its discontinuity is a conspicuous feature of the Ruahine Range, with tongues of

26 Transactions – BOTANY VOL. 3
tussock and leatherwood breaking its outline, mainly down gullies which, however, show traces of a former tree canopy not only in fallen logs but also in dead standing trees whose branches may still outline a former wind roof. There is also similar widespread evidence of a former timber line some 200-300ft above the present one (Plate 4A). Ring counts of mountain beech in the Ruahine Range have not produced a figure of above 150 for the largest trees, and, on the assumption that few dead trees will remain standing for more than a further 50 years, this gives evidence of a considerable change having taken place over as recent a period as the last 200 years. (It must be noted that these ring counts were made in the field. Wardle (1963b, p. 35) gives consistently higher counts and considers (pers. comm.) that scanning by microscope is necessary for slow-growing timber.) Not only has there been a retreat in altitude, but the pattern of the slopes, with trees clustered on the drier ridges and knolls and regeneration confined to them, with leatherwood and tussock replacing them in gullies, suggests that conditions have become wetter.
A timber line community of old gnarled mountain beech over a closed tussock floor of Chionochloa conspicua ssp. cunninghamii and/or snowgrass is associated with level or slightly sloping ground and so has presumably some connection with drainage (Plate 4B). There is little sign of replacement of the canopy, which may be so open as to approach a park-like pattern of separate trees. This is described more fully in dealing with its occurrence in beech-cedar ecotones (p. 38).
A terrace above the Waikamaka valley at an altitude of 3,600-4,000ft (that is, towards the lower limit of mountain beech) though of small extent is of particular interest; for when first observed in 1940 it had the same pattern of dead trees standing in Chionochloa conspicua meadow, but has since changed with some rapidity. Aerial photographs taken in 1950 showed an unusual texture of scattered conical trees appearing, and by 1954 in addition to these the tussock had been largely replaced by dense scrub while most of the dead trees had fallen. The conical trees were found to be, rather unexpectedly, mountain beech some 12-15ft in height and 33-34 years of age, the scrub mainly species of Coprosma but with some leatherwood and tussock, the latter tending to dominate up the slope. A local fire dating prior to 1922 would seem to be the most likely origin of this pattern, but no trace of charring has been found.
In the snowgrass above the present timber line what appear to be scattered buses of mountain beech may occur among the logs of the former timber line, particularly along the divide above the head of the Maropea River, but still more so on lower ridges east of the divide, as for instance the Shut Eye ridge, which is particularly exposed to violent downdraughts and eddies in westerly gales. On examination these are often found to be the branches of prostrate trees whose trunks may be up to 20ft in length and a foot in diameter.
On the western and southern boundaries of the Central Ruahine area where mountain beech is replaced by cedar forest and scrub, several distinctive patterns occur which indicate the downward and outward movement of beech forest into the neighbouring formations. This is described in more detail at the end of the Western Ruahine section where the cedar-beech ecotones are discussed.
CENTRAL: CEDAR – DACRYDIUM BIFORME
Cedar, more or less associated with Dacrydium, occurs over most of the range m mountain-beech forest, most commonly at an altitude a little below 4,000ft, where its corneal heads frequently form a distinct belt. It is, however, almost impossible to find a sound tree, even apparently healthy and vigorous trees commonly having only an inch or so of sound timber, with close and even rings, surrounding a core of rotten wood. Seedlings or saplings are practically non-existent

NO. 3 ELDER – Vegetation of the Ruahine Range 27
in the upper part of its range (see Text-fig. 2), but become more frequent lower down, and where cedar occurs below its usual range they may become common. One all-aged stand occupies a limited area on a scrubby shingle slope on the northern boundary towards the foot of Pohatuhaha (c. 3,300ft). Narrow spurs above the inner river valleys frequently carry a conspicuous line of cedars also descending below their usual range.
A pattern of scattered bushy Dacrydium biforme on the tussock spurs that rise above the beech timber line is frequently seen throughout the Central area.
CENTRAL: UPPER STREAM BANK
Mountain beech commonly colonizes river terraces and stream banks well below its usual range. In the lower part of the Waikamaka valley silver beech, extending from the adjacent Mokai Patea colonies (Western Ruahine), is common and may even be co-dominant with mountain beech.
A characteristic shrub of the upper river terraces near the timber line is Hebe truncatula, which is also a pioneer colonizer of screes and is described in that connection. Coprosma rugosa is abundant in many places, though less widespread, and Muehlenbeckia axillaris is a conspicuous mat plant of these higher reaches, where the proportion of subalpine plants of the same form increases with altitude.
CENTRAL: SUBALPINE
Except for the major volcanoes all the mountains of the North Island lie well within the range of vegetation, but a considerable proportion, exceeding 30%, of the area of the Central Ruahine lies above the timber line. Dissection has not gone as far as on the Tararua Range, so that there is a mixture of narrow ridges and wide rolling summits with a considerable proportion of bare rock and scree on their sides. Two closed communities cover much of the surface; leatherwood scrub dominated by Olearia colensoi below 4,700ft; and snowgrass tussock, which reaches the highest altitudes and covers considerable areas, particularly on the western side. Extensive peat bogs occupy the upper part of the Mokai Patea plateau, and similar smaller bogs occur north of Tupari and on the Hikurangi Range in the lee of the higher crests. There are also small boggy areas on ledges where snowdrifts accumulate; these are occupied by a distinct community of mat species. Screes in the initial stages of stabilization carry a sparse vegetation of specialized plants with extensive root systems.
CENTRAL: SUBALPINE SCRUB
Olearia colensoi and Senecio elaeagnifolius are present in and above the timber line throughout the Central Ruahine, but typical dense leatherwood scrub dominated by the former is only encountered in limited areas, mainly in basins or on the broader spurs below 4,700ft (List 7).
On the southern boundary, in the near absence of mountain beech, the steep south-facing slope into the Oroua River is occupied by a major scrub formation with a vertical range approaching 1,000ft. (This was the scene, in 1948, of the arduous Howlett’s search for a crashed RNZAF plane which had disintegrated in 10-foot leatherwood.) This adjoins the induced scrub-tussock association of the extensive Daphne Ridge burn of the 1880s, where the original pattern is obscured.

28 Transactions – BOTANY VOL. 3
Over most of the Central Ruahine a characteristic scrub-tussock formation extends downwards into the numerous gaps in the timber line, Chionochloa conspicua replacing snowgrass below 4,500ft. Olearia colensoi has not been found on the Kaweka or Kaimanawa Ranges to the north and Senecio elaeagnifolius occurs only sporadically, so that leatherwood scrub as a formation is absent from a belt 40-50 miles wide. On the Ruahine Range the northernmost occurrences of Olearia lie on an oblique line from the lower end of the Mokai Patea plateau to the vicinity of Apias Creek on the NE plateau, thus roughly following the boundary between the western and central areas. The distinctive features of leatherwood along this boundary are described more fully in the western Ruahine section of this paper.
Though scrub as a formation ceases at about 4,700ft, Olearia colensoi persists as a sprawling shrub, often only a few inches high, to above 5,000ft, typically on south-facing slopes. Dracophyllum recurvum has a somewhat similar range and habit, but several woody shrubs, the chief of which are Senecio bidwillii, Hebe odora, and Hebe subsimilis, maintain an erect bushy form scattered through snowgrass tussock to above 5,500ft (List 5B).
A comparison of earlier species lists with more recent ones shows that considerable changes have taken place over the past 20-25 years in the composition of alpine scrub; some, once regularly listed as common and occasionally even as abundant, are now inconspicuous or recorded only on inaccessible sites. All are species favoured by deer, and the period covers the build-up of the deer population along the range. Neopanax colensoi has almost completely vanished; Aristotelia fruticosa (the Tararua variety) and Olearia arborescens have become infrequent, as has Gaultheria subcorymbosa south of the Waipawa Saddle. Until recently this last species had been listed as absent north of the Saddle but the recent sighting of a solitary specimen on a cliff in the Mangatera, together with the consistent rarity of Aristotelia in the northern half of the range, suggests that these had already been reduced by browsing before the main southward movement of deer began.
In addition to the species mentioned, Coprosma foetidissima has been almost invariably hard-browsed, though survivors are frequent. Its recovery in areas of high-priority hunting such as the Waipawa headwaters is now becoming conspicuous. Under the scrub Polystichum vestitum, once abundant and Ranunculus insignis, formerly luxuriant along watercourses, only survive reduced in number and size.
The main subalpine scrub species are listed downwards in the altitudinal sequence in which they occur (List 7). Others of minor importance are Gaultheria subcorymbosa and Pimelea buxifolia south of the Waipawa Saddle the Hebe truncatula group where scrub is encroaching on to scree, and of very local occurrence Dacrydium bidwillii, though this is more characteristic in snowgrass in the neighbourhood of bogs. The ball koromikos H. odora and H. venustula and the whipcord H. subsimilis only enter the margins in stunted scrub and are hardly typical scrub species.
CENTRAL: SUBALPINE TUSSOCK
Above the timber line the most widespread formation is tussock grassland dominated by a uniform, rather narrow-leaved form of snowgrass, Chionochloa pallens, distinct from either of the two species that occur in the Tararua Range. Where the divide is narrowest tussock mainly occurs on the easier western slopes, but where the crest broadens out to north and south, and particularly on the

No. 3 ELDER – Vegetation of the Ruahine Range 29
wide summits of the Hikurangi Range and Mokai Patea plateau, it covers extensive areas. At lower altitudes on the Mokai Patea snowgrass is replaced on wetter ground by red tussock below 4,500ft. Taramea (Aciphylla colensoi), formerly associated with snowgrass in sufficient abundance for its dagger-like leaves to be mentioned by Colenso as a considerable obstacle to his lightly clad Maoris, and giving its name, Puketaramea, to a peak on his route, has since become comparatively rare. It is obviously keenly relished by browsing animals -certainly by deer and perhaps by hares – and over the past 30 years has become confined to inaccessible sites except in one or two limited localities. There is evidence of its recovery in the past 4-5 years, presumably connected with a decline in deer population, accelerated towards the eastern side of the range by intensive hunting.
Low-growing shrubs are scattered through the tussock, and a number of low-growing plants, most of them herbaceous, are plentiful beneath them (List 5A). On more exposed sites, tussocks are more widely spaced and less luxuriant, allowing low-growing plants such as Celmisia spp. to appear in greater variety and to form a larger proportion of the ground cover. In still more exposed sites fell field appears: a surface of lichened rock fragments carrying an open community consisting partly of plants associated with snowgrass (which commonly persists in scattered stunted tussocks), but with the addition of certain plants more or less confined to that situation (List 5c).
CENTRAL: BOG
The main peat bogs lie on the western and northern plateaux and share in different degrees elements of a common flora. This has been most fully recorded in the Northern Ruahine and can best be dealt with in the account of that area. The upper part of the Mokai Patea is the most extensive area of peat bog lying outside the Northern Ruahine and within the Central Ruahine area as here defined.
Floristically this bog is most closely linked with the Northern Ruahine bogs and to a lesser degree with the western (Whanahuia) bogs (List 9).
Smaller bogs occur on the divide near Tupari, south of Te Atua Mahuru, and on the western end of the Hikurangi Range. In each case tarns with raised sphagnum rims or their relics are evident, but otherwise the associated plants in most cases are merely those of wetter sites in the neighbourhood. Near Tupari, however, a somewhat larger bog area (perhaps the traditional waterhole Ngaroro a Kahupakiri) carries a number of plants (for example, Drosera arcturi, Myriophyllum propinquum, Hypolaena lateriflora, and Geum leiospermum) which do not occur over a considerable area around but are characteristic of the larger bogs.
On the Mokai Patea peat formation is not now active except in one or two areas round tarns, and the erosion of peat or the invasion of its surface by tussock vegetation is characteristic (Moar). At higher elevations snowgrass is the tussock species, but below 4,500ft red tussock is increasingly prominent on wetter ground with snowgrass confined to drier sites. Buried timber has been observed in the vicinity up to nearly 4,700ft but not in the bogs themselves, and as far as investigation has gone it only occurs intermittently.

30 Transactions – BOTANY VOL. 3
CENTRAL: SCREE
Screes, rock falls and shingle-choked watercourses are conspicuous through most of the Central Ruahine, particularly along the eastern fault scarps, an impressive example being shown in the aerial photograph of the Waipawa Saddle (Plate 1), where the contrast between the tussock-covered western slopes at the head of the Waikamaka and the great area of rock faces and shingle on the Hawke’s Bay side is strongly marked. There is also a considerable development of screes in some of the inner valleys, particularly those of the Makaroro, Mangatera, Kawhatau, and Pourangaki. A scree system in the northern head of the Mangatera, is closely associated with a series of scarplets and on a small scale shows the most conspicuous relationship of faulting to scree formation in the range.
Where the whole face of the range can be viewed, as for example the Hikurangi Range from across the Kawhatau valley, or Te Atua Mahuru from Remutupo, it is clear that there is some sequence of scree formation. A former system or systems of screes now completely revegetated in snowgrass or scrub has interspersed with it, rather than superimposed on it, a comparatively recent active system. The general impression suggests two or more cycles of scree formation. Elsewhere individual screes occur in varying stages of stabilization, which suggests that scree formation can be a continuing process, each scree reaching maturity when its head has eaten back to the crest of the ridge. At this point, where the slope eases out to a surface of fine rubble or clayey soil, a scattered community (see List 6A) is conspicuous.
Further down the slope characteristic scree species with wide-ranging root systems frequently occur in running shingle (List 6B)
As the base of a scree becomes stable it is invaded by Hebe truncatula, a pioneer species of such sites m the Central Ruahine, where it forms characteristic pure sands whose dense canopy opens up after a period of about 20 years. Though listed as a true species individual plants show variations, mainly in leaf shape, texture, and colour; in cultivation, when it is grown from seed, two flowering periods (November and February) are common, while some bushes consistently produce up to 80% of branched racemes. These characters suggest a hybrid origin, but the parentage is in doubt as the geographical and taxonomical considerations do not agree.
The process of recolonization has been followed on a 1,100ft scree opposite the Waikamaka Hut. It shows on a 1933 photograph as apparently bare shingle, and in 1939 was still free running shingle over almost the whole of its length. The Hebe then began to appear at the foot and inward from the flanks; subalpine plants and snowgrass started to move in higher up, and, although the scree had not yet eaten back to the crest and still had an overhanging lip corniced in winter scattered prostrate plants began to establish footing across the upper surface. This is now approaching a closed cover, while the lower third has become an almost impenetrable Hebe thicket. Between them a few lanes of loose shingle persist on which scree plants are becoming established, Epilobium pycnostachyum within the last five years or so.
One interesting pattern of scree occurs on the back of the Shut Eye Ridge facing the northern head of the Waipawa, where older screes are occupied by 50- to 60-year-old mountain beech and are now being undercut by a recent active scree. Charred timber has been found on either side of the valley, so it is possible that the beech regeneration and subsequent erosion are effects of a fire occurring about 1900.

No. 3 ELDER – Vegetation of the Ruahine Range 31
NORTHERN RUAHINE (Map 3)
The Northern Ruahine consists of three blocks – the NW and NE plateaux, and the Pohokura basin between them – each of which has botanical characteristics of its own. The NW plateau is a southerly extension of the Ngamatea plateau and its affinities are with the Inland Patea. Both the Pohokura basin and the NE plateau have closer floristic relations with Hawke’s Bay, so that the Northern Ruahine could be divided into NW and NE areas, though it is here treated as a whole. Several species with a northern distribution extend on to it, more especially on lo the NW corner, and a number of coastal species persist in the eastern gorges, while several species that are prominent further south drop out or become less prominent (e.g., leatherwood, and red beech).
Both plateaux at the time of European occupation were occupied by a mosaic of mountain beech and red tussock like the Inland Patea and Ngamatea country to the north. It is still a matter for speculation as to how far these were Maori fire patterns, as the evidence has been so generally overridden by European burning
The Pohokura basin between them, however, when first recorded, carried what was evidently an induced cover of tall kanuka scrub with islands of matai-dominant forest; it may be matched with the Omahaki basin downstream of it on the Kaweka Blowhard
One difference between the Inland Patea and Ruahine plateaux is the sharp falling off of the Taupo ash to a thickness of less than an inch, and associated with this the absence of subsurface charcoal. At the same time there is evidence (cf. p. 34) particularly in bogs, of the replacement of Dacrydium biforme by red tussock at about the level of the Taupo rush.
Almost the whole of the red-tussock grassland has been stocked with sheep, and for the most part burnt, over a period of up to 80 years, and cattle have been run on parts of it, though perhaps more recently. It was also the first part of the range to be heavily browsed by deer, so that both tussock and forest have been considerably modified from their pre-European state. Evidence of earlier Maori burning is still indefinite and undated, but seems likely, as it accumulates, to be recognized as important.
NORTHERN: PODOCARP FOREST
Vestiges of podocarp forest remain at several points round the margins of the area and in the Pohokura basin, generally at rather low altitudes, beech forest generally dominating above 2,000ft. The exception is an isolated area of podocarp-hardwood forest towards the head of the Wild Sheep Spur at nearly 3,000ft, between the Ikawetea and Makirikiri Streams. These remnants resemble those of the Kaweka Blowhard and differ from the rimu-dominant stands common elsewhere along the margins of the range in that matai is the dominant species, quite often associated with totara, though rimu is a fairly constant minor component. Kahikatea occurs frequently, but on the other hand miro is rather rare. With podocarp forest are associated such broad-leaved species as black and white maire. Mountain maire, though rare, occurs at Herrick’s Hut (1,500ft).
NORTHERN: RIVER CLIFF
In the gorges of the Ngaruroro, Taruarau, and Rangitikei Rivers at the lowest altitudes in the area (ca. 1,000ft) there occur a number of pockets of strictly lowland species which also occur at the other end of the range in the Pohangina and

32 Transactions – BOTANY VOL. 3
Manawatu valleys. Myoporum laitum, Dodonaea viscosa, Rubus squarrosus, and the rupestral form of Hebe angustifolia var. parviflora are characteristic. The north-eastern communities have in addition a striking Hawke’s Bay coastal component (List 10) mixed with such Inland Patea endemics as Hebe colensoi and Myosotis eximia. There are also several miscellaneous and more localized species: Coprosma virescens and C. rubra (Rangitikei and Pohokura) an extensive colony of Solanum aviculare var. albiflorum (Big Hill Stream) and Hebe angustifolia var. arborea.
NORTHERN: RED BEECH FOREST
Except in the upper Ikawatea valley and in one or two stands of limited extent, as for example at Toimaru, red beech becomes infrequent and is absent from most of the area, its place being taken by mountain beech, and in the Paramaho Bush by black beech merging into a black-mountain swarm. The latter is a feature of some interest, as the Paramaho Bush lies in a zone several miles wide which extends westwards into the SW Kaimanawa Range and also makes its appearance in the opposite direction-to the NE on the Kaweka Blowhard, in which a black-mountain swarm is dominant in the relics of forest that remain. Elsewhere in the Ruahine Range, where black and mountain beech occupy different altitudinal belts separated by the red-beech belt, the two species are recognizably distinct, but where the red-beech belt thins out (as in the Makaroro mill workings which lie just outside the boundary of the northern area) with black and mountain beech present on either side of it, a narrow belt of hybrids occurs in the red beech along the 2,000ft contour.
Evidence has recently come to light from buried beech leaves on the margin of the Taupo shower in the Kaweka Range that there has been a change within the last 2,000 years in the composition of a black-mountain swarm (Cunningham).
NORTHERN: MOUNTAIN BEECH FOREST
Few points in the area exceed 4,500ft, so that practically the whole lies within the altitudinal range of mountain beech. However, red tussock dominates the wider crests and easier slopes, with beech forest dominating the steeper slopes and narrower ridges. Tongues of forest or occasional clumps of trees may occur in the tussock areas, and narrow lines of wind-blown trees across the tussock slopes may persist and are characteristic (Plate 5B). These narrow lines appear to mark a change of slope or the margin of deeper peat. Thickets of stunted beech and Dacrydium biforme also huddle in troughs close to the highest point of the No Man’s bog (4,500ft), and here shelter from wind is an obvious factor of their persistence as well as depth of peat. The troughs appear to be due to the checking of Sphagnum formation under the dense canopy. Where the formation is more open the shelter of individual bushes favours Sphagnum, and mountain beech in this situation largely roots in it. Dacrydium biforme, however, tends to supplant beech where the drainage is poor. Cursory observations on No Man’s Land suggest about 1 ½ ft depth of peat as the limit for beech and something more than 3ft for Dacrydium. Along much of the level surface of the plateau an open type of beech/scrub forest occurs, with scattered crowns and a high proportion of dead trees over a dense divaricating scrub of browse-resistant or unpalatable species. The earliest concentration of red deer occurred here and they must largely be held responsible for this pattern, but there is also a longer-term cause. Clusters of dead beech trees tend to occur on waterlogged ground on which Dacrydium biforme is coming in; the entry of this species indicates an early stage of formation of peat bog.
No. 3 ELDER – Vegetation of the Ruahine Range 33
Where forest occurs on the slopes the canopy is usually closed, but until recently undergrowth was sparse and on the eastern side almost completely absent even before goats came in from the Wakarara Range. Young beech regeneration is conspicuous at the present day along the brow of the plateau; but it is absent both from the crest and from the slopes under dense canopy. The relation of this to drainage and light is clear; its relation to browsing pressure is dealt with later (Appendix 1).
At the north-eastern extremity of the range cedar is associated with beech, but at lower levels and hence mainly on the slopes. It is not in great quantity and appears to be dying out, as dead trees are frequent in forest. In scrub, how ever, on the Pohatuhaha Spur at 3,300ft there is one notably vigorous all-aged stand, while occasional saplings occur at lower levels along the foot of the Pohatuhaha Range. A somewhat similar pattern of failure above and success below has been noted towards the other end of its range at Umutoi (Western Ruahine).
NORTHERN: TUSSOCK
The dominance of red tussock is associated with level or gently sloping ground and hence frequently with the formation of peat, certainly with a reduced rate of drainage and presumably with a higher soil acidity. Where snowgrass is associated with red tussock it shows a marked preference for drier sites, as on mounds of drift pumice (Ohawai). A tendency has also been noted along forest margins (Ruahine Comer) for snowgrass to replace red tussock.
As practically all areas of the Northern Ruahine carrying red tussock have been utilized for grazing accompanied by periodical burning, considerable modification has taken place. Where stocking and burning are still carried on introduced plants have appeared, with areas in which scab-weed communities have replaced tussock, extensively so near Makirikiri trig on the rim of the limestone scarp of the NW plateau.
On the NE plateau the tussock and topsoil have been removed from two or three localities, exposing buried timber in sufficient quantity to make it evident that the wide tussock slopes of Ohawai once carried forest. Both here and at Apias Creek, where the stumps are exposed in situ, this was predominantly Dacrydium (mostly D. biforme) forest, which at Apias was rooted above a horizon of weathered ash and a few inches below a Taupo ash horizon (Plate 5A). The Piopio exposures south of the plateau are in snowgrass but have also uncovered wood, in this case almost certainly Olearia colensoi. These exposures could have been initiated by the trampling of sheep, perhaps on a surface exposed by the burning of tussock, or later by the burrowing of rabbits or even later still by the trampling of deer. They have certainly developed through the action of wind on Ohawai (where drift dunes colonized by snowgrass have developed on their margins) as well as of water, the latter being more evident at Apias.
NORTHERN: BOG
The Reporoa and No Man’s bogs on the NW and NE plateaux respectively carry the largest and most specialized flora and the bogs farther south along the range show some relationship to them, as has already been mentioned in dealing with those on the Mokai Patea and near Tupari (List 9). Features common to all of them, though most developed in the north, are the shallow tarns with raised sphagnum rims, the association with red tussock, which may even mask the existence of peat, and the presence of ash horizons.

34 Transactions – BOTANY VOL. 3
Reporoa Bog. This bog lies on the Mangaohane limestone at an altitude of 3,800ft and has been the most closely investigated botanically. Of 119 species listed by Druce in 1948, 13 were not previously recorded north of Cook Strait, and a further 33 had been recorded in the North Island only from the Tongariro National Park.
Peat profiles show a depth of 16ft and the main features of their pollen are the steady increase from the base of the beech as against podocarp pollens, the prominence of rimu as against all other podocarps, and the extreme scarcity of cedar.
No Man’s Bog. This bog lies at an altitude of 4,500-4,300ft about the timber line and several hundred feet above Reporoa. Red tussock covers most of its surface, with a little gnarled and frequently moribund Dacrydium biforme and mountain beech where the peat is shallower. The blanket of peat is generally 5-6ft deep, its chief features being the clearly defined ash horizons and the abundance of preserved timber (Plate 6A). It has been found possible with a sounding rod to distinguish not only logs but layers of twigs and even volcanic ash to depths exceeding 5ft. By this method some 200 probes have been made on a grid covering about an acre which show a fairly even distribution of timber with a possible maximum a little below the Waimihia ash over a period of several thousand years (that is, from more than twice the depth of the recognizable Waimihia ash shower) until about the time of the Taupo shower (1840 years B.P.) after which there is a pronounced diminution in the amount of timber.
Samples of timber have been collected to a depth of 30in, identified by wood structure, and a selection of them carbon dated (Grant-Taylor and Rafter 1962). Except near the bog margins all timbers have been identified as Dacrydium and most as D. biforme.
A plot of C14 ages against depth suggests that the rate of peat accumulation had been decreasing for a long period prior to the Taupo shower, followed by an increase (Plate 6B). The progressive disappearance of the earlier ash showers towards the margins shows that this bog has been expanding laterally for several thousand years.
The present distribution of Dacrydium biforme on the NE plateau shows that it will flourish on deeper peats than mountain beech will tolerate. Buried Dacrydium logs with a maximum of at least 4ft of peat below them are widespread in the No Man’s bog and, as the frequency of timber falls off sharply above that level, this suggests at first sight that the depth of peat is the limiting factor. Against this is the abundance of subsurface timber in the form of stumps in situ on clay subsoil as well as logs further along the plateau (Ohawai and Apias). The presence of the Taupo ash horizon in all these exposures shows that the replacement of timber by red tussock on these different kinds of sites was approximately contemporaneous. Some indirect association with the Taupo eruption is a possibility, as for example fires spreading on the periphery of the area to the north where hot ash destroyed the vegetation.
Pollen analysis shows a dominance of beech over the whole period of peat formation, with a general decrease of Dacrydium biforme type pollen prior to the Taupo shower and only intermittent appearance since. The profiles from No Man’s do not match those from the western (Mokai Patea and Whanahuia) bogs, but no detailed comparison has yet been possible (N. T. Moar, pers. comm.).

No. 3 ELDER – Vegetation of the Ruahine Range 35
NORTHERN: SCRUB
As a result of stocking and burning of the more open country and prolonged browsing of the whole area by deer, there is perhaps no natural scrub formation left in the area. Forest undergowth [undergrowth] (List 8) appears to have developed into a dense, divaricating scrub form, particularly where mountain beech is dying out and ceasing to form a canopy. Olearia colensoi only occurs towards the southern boundary; seedlings of Senecio elaeagnifolius are remarkably widespread on protected sites, but the rarity of adult plants suggests that this species has been reduced by browsing.
Towards the NE extremity of the range fire-induced manuka scrub is extensive up to about 3,500ft. This resembles the scrub of the Kaweka Blowhard in species composition and relative abundance of Leptospermum ericoides. Its heavy defoliation, presumably by opossums, behind Kereru is a recent (1962), and on the steep slopes, an unwelcome development (List 11A).
An open scrub community on higher exposed crests, particularly the Otupae Range and Te Koau Ridge, also shows affinities with that of similar situations in the Kaweka Range. Manuka takes the same prostrate form, but the frequency of clumps of Griselinia littoralis huddled in the lee of small hummocks is peculiar, especially in the presence of deer (List 11 B).
NORTHERN: LIMESTONE CLIFF
Besides the river-cliff communities which have been described in an earlier section several limestone blocks (Aorangi, Rakaunui a Kura, and Potae) exceeding 4,000ft in height, which are striking features along the southern margin of the NW plateau, support a distinct assemblage of plants on their cliffs and in the numerous clefts of the plateau itself (List 12).
WESTERN RUAHINE (Maps 3 and 4)
The clear-cut line of cedar forest across the end of the Mangaohane tussock marks the northern margin of the Western Ruahine forest. Cedar forest occupies most of the Mangatera basin and runs south along the outer flank of the range till the Oroua River emerges, then continues in an increasingly modified form as far as the mouth of the Pohangina Gorge, where it is indistinguishable from the forest of the Southern Ruahine in which cedar is no longer dominant.
Although cedar occurs throughout the range, as does Dacrydium biforme, it is only where mountain beech is absent that one or the other species can dominate the timber line so that on this basis the boundary between Central and Western Ruahine follow the edge of the mountain-beech forest. This boundary begins near the divide at Potae and runs obliquely SW across the Mangatera valley to the Mokai Patea. A narrow strip of cedar forest continues south along the western face as far as the Whanahuia Range. Cedar continues to dominate on the outer face of this but on its inner side Dacrydium biforme becomes increasingly important in a wide belt running obliquely SE across the Oroua and Pohangina valleys to the Hawke’s Bay side of the range.
As earlier stated, the upper Pohangina valley could be treated as a separate natural area. but for simplicity’s sake is here considered as an extension of the Western Ruahine area which then is distinguished by the absence of mountain beech, the presence or red beech, and the dominance of cedar and/or Dacrydium biforme at the timber line.
The Western Ruahine thus consists of two main areas, a triangular one in the north and an oblique band across the range in the south, joined by a narrow outer strip across the foot of the Mokai Patea tussock.

36 Transactions – BOTANY VOL. 3
WESTERN: PODOCARP FOREST
Podocarp forest has generally been cleared and milled along the western flank of the range. Such vestiges as remain are mainly rimu scattered through redbeech forest, but this may not give a true indication of the composition of the virgin podocarp forest. In the two or three unaltered areas of podocarp forest of any size that remain – on the western slope of Aorangi and in the Pourangaki valley, to which may perhaps be added a third in the Oroua valley – rimu is not generally the dominant podocarp.
The northern forest below Aorangi is strikingly distinct. Here matai and totara are co-dominant, with white maire (Gymnelaea (Olea) lanceolata) plentiful as a sub-canopy species. Rimu only appears infrequently at the upper extremity of the belt, and other podocarps are absent. A fair amount of light reaches the floor, which is occupied by an open undergrowth of an unusual assemblage of small trees and shrubs. Grasses, notably Microlaena polynoda, form a dominant floor cover (List 13).
In the southern area, though rimu is plentiful, matai tends to dominate, and associated with them are miro (Podocarpus ferrugineus) and to some degree kahikatea (P. dacrydioides). White maire, as elsewhere, is commonly associated with podocarps, red beech is a component at higher altitudes, and below 2,000ft, totara, rewarewa (Knightia excelsa) and rata.
WESTERN: RED BEECH FOREST
Over most of the area red beech dominates an altitudinal belt above podocarp forest to a mean of 3,600ft, corresponding to its distribution elsewhere. At either extremity of the area, however, red-beech forest as a formation breaks up; north of the Mangatera red beech persists up to its Otorere tributary, but is elsewhere replaced by cedar forest, in which occasional stands of red beech can be recognized, generally at rather low altitudes (below 3,000ft), which from their close canopy appear to be young growth in process of invading deteriorating cedar forest. The northernmost stand seen on this side of the range occupies a sheltered gully near Racecourse Flat (Akuratawhiti) on the Rangitikei River about 2,000ft above sea level in the extreme NW corner of the area.
Towards the south kamahi is increasingly associated with red beech as a sub-canopy species, and eventually replaces it. The sequence along the outer flank of the Ngamoko Range is clearly marked. Red beech is dominant south of the Oroua valley as far as Coal Creek, beyond which it rapidly becomes infrequent in the upper part of its range, though it continues to dominate on the lower slopes interspersed and to some extent intermingled with podocarp forest. At the mouth of the Pohangina gorge kamahi is practically the sole dominant of the belt above the podocarp forest and the pattern is now that of the southern Ruahine.
WESTERN: CEDAR-DACRYDIUM FOREST
Where cedar is dominant at the timber line its conical heads make it conspicuous both on the ground and in aerial photographs. Red-beech forest has a maximum altitude not much lower than that of cedar so that where the two species are associated the narrow belt of cedar forms a distinctive band between the beech below and the leatherwood above.

No. 3 ELDER – Vegetation of the Ruahine Range 37
Towards the north where red beech is absent, as in the Waipato basin between Aorangi and Titapu, cedar occupies a wide altitudinal range extensive in area. But the high proportion of dead crowns, the open canopy, and on the ground the conglomeration of fallen logs and bush lawyer are evidence of a considerable change in progress, with some indication, as previously noted, of red beech invading at lower levels, and in the neighbouring Otorere, of mountain beech at higher levels. A striking exception to this deterioration of cedar forest is noticeable in aerial photographs, on which limestone blocks such as Aorangi and Rakaunuiakura, as well as less obvious ones, can readily be picked out by the density of stands of cedar and Dacrydium biforme (Plate 7A). Cedar-dominant forest occurs intermittently NW of the Ruahines along the margin of the Inland Patea tussock and round the western side of the volcanic plateau to at least as far north as the edge of the Waimarino Plains. The vigour of cedar forest on this arc, in the Kaikawaka Bush, on the limestone scarp of the SW Kaimanawa, and perhaps on the Tertiary scarps further west is supporting evidence of its affinity for limestone.
South of the Mangatera River to and including the Mokai Patea, burns, pre European as well as European, are at least partly responsible for discontinuous forest and complicated patterns dominated by cedar, mountain beech and, in limited areas, Dacrydium biforme, with the addition of small colonies of silver beech. This last is absent from the rest of the range and indeed from the whole Rangitikei catchment upstream of the Moawhango confluence. On the lower part of the plateau small areas of cedar saplings, though browsed by stock or deer, have established themselves in tussock, presumably after European burning. (Similar browsed cedar invading tussock after fire has been seen in the southern Kaimanawa Range.)
Generally cedar tends to dominate in a narrow outer strip along the Mokai Patea and the Hikurangi Range, but it is noticeable that a cedar timber line persists for some distance up the valleys on the west- or south-west-facing slopes of the Kawhatau and Pourangaki.
Farther south again Leatherwood scrub becomes increasingly extensive, and an increasing proportion of Dacrydium biforme occupies the timber line except on the outer face of the Whanahuia Range, where cedar dominates. Dacrydium and cedar are approximately co-dominant on its inner face, but this is not obvious because the broken crowns of cedar on exposed sites cannot readily be distinguished from those of Dacrydium. As further east again in the upper Pohangina valley, the ridge crests are distinguished by the smooth wind roof of Dacrydium, while cedar in its characteristic form appears only infrequently in the red-beech forest below. Tolerance of wind appears to be an important distinguishing factor between the two species.
As the southern boundary is approached the two species tend to separate out, cedar to the lower levels, where it is conspicuous, keeping its conical form, on ridges and knolls, Cedar Knob on Shorts’ Track being a typical example. Dacrydium on the other hand, maintains its altitude, but in a stunted form, level with the surrounding scrub along exposed ridges, conspicuous in aerial photographs as darker ribbons in the leatherwood. Some completely misshapen cedar merges with it. There is thus no distinct timber line, kamahi forest also becoming stunted and continuous with the scrub canopy. These patterns continue into the Southern Ruahine, where they are characteristic.

38 Transactions – BOTANY VOL. 3
WESTERN: CEDAR-BEECH ECONTONES
The Western (cedar) timber line is considerably below the Central (mountain beech) timber line, and the transition zone from one to the other shows several interesting features. In general the mountain-beech timber line descends to the level of the cedar belt, which is more or less continuous throughout the beech forest itself, but at the junction point pure stands of Dacrydium biforme are conspicuous, particularly capping exposed spurs, on which they replace cedar.
Some of these Dacrydium stands appear on aerial photographs to be of considerable extent, as for example on the western side of the Mokai Patea and in the head of the Oroua. There is no obvious explanation for their presence. The dominance of Dacrydium on exposed and better drained sites here, and in the Central Ruahine generally, contrasts with its tolerance of boggy sites in the Northern Ruahine.
As the mountain-beech timber line tapers off into Dacrydium/cedar it tends to open out into a pattern of large beeches dotted about in tussock, giving almost a park-like appearance which is particularly noticeable on easy slopes and therefore probably dependent on drainage. Though these are over-aged trees with no replacement in sight the suggestion has been made (Wardle, pers. comm.) that this stage does not represent a general retreat of the timber line, but is a wide spread formation which until the arrival of deer had sufficient regeneration to maintain its open pattern.
The narrow cedar belt above red-beech forest has already been described. In the vicinity of mountain-beech forest the persistence of scattered trees of mountain beech between the cedar and red beech is characteristic of this formation.
In the most extreme case noted, down the Oroua valley, a discontinuous band of mountain beech is still encountered between cedar and red-beech forest some three miles south of the last of the mountain-beech timber line.
Most of the Mokai Patea timber lines have been affected by fires of various dates. There is, however, one compact area at the head of the Makino Stream which presents in miniature most of the patterns of the ecotone which have been observed elsewhere, so that it may be assumed to have escaped fire and is described as typical to a considerable degree of the ecotone, though not of the immediate neighbourhood. Above the head of the valley the marginal outlier of mountain beech slopes downwards from a maximum of 4,300ft till it meets a horizontal belt of cedar. This does not extend as far as the head of the valley, but in the upper part three small spurs at that level are capped by close clumps of Dacrydium biforme, which also occurs immediately above the cedar, and Phyllocladus alpinus is plentiful in and above it. In the vicinity large bushes of Olearia colensoi are scattered through the snowgrass meadow below Colenso Trig.
Below the cedar belt, which is narrow, red beech is dominant down the Makino valley, but the intermediate belt here is occupied by a mixture of all-aged silver and mountain beech in proportions ranging from 3:1 to 1:1. The predominance of silver beech in this situation is a purely local peculiarity. Though silver beech occurs at several points on the plateau and well up the Waikamaka valley to the east of it, mountain beech is dominant generally speaking
Piringa (Plate 2). This is an outlier of cedar timber line, and though it is well inside the Central Ruahine area it has affinities with the central-western ecotone, so that it can most conveniently be described here. It is of particular interest as it forms the lower margin of a bald, similar to those in the Appalachian Mountains, described by Mark. This occurs on the northern (i.e., sunny) face of Piringa, a peak (4,300ft) above the upper fork of the Maropea River.

No. 3 ELDER – Vegetation of the Ruahine Range 39
It consists of a belt of snowgrass and scrub separating two forest types, mountain beech above and cedar below, the latter at 3,800ft, an altitude approaching the upper limit of cedar and rather above that of red beech but near the lower limit of both snowgrass and mountain beech. It thus lies close to the limits of several dominants. The slope is steep (about 30°); the eastern end is in continuous snowgrass tussock, the western mainly in scrub which runs out, on its downhill side, on to bare shingle slopes approaching scree. Where soil persists it is 2-3ft deep with charred timber appearing frequently 12in below the surface. This matches the sub-surface charcoal found to the south across the Waikamaka valley on the prominent tussock spur that runs off the Mokai Patea, and thus links it with the legendary Tahuarongotea fire. In the opposite direction the tussock crest of Puketaramea may have had the same history, and the curious scrub face which carries the name Otukota (p. 18).
The open slope of Piringa is much favoured by deer, which may be presumed to have reached their peak of population in this part of the range about 25 years ago and to have modified it considerably by browsing and trampling on the steep slope.
The crest of the ridge and its upper slope is occupied by mountain-beech forest with a high proportion of pole timber and indication of a vigorous spread downwards on to the bald until a comparatively recent period when, to judge by sapling gaps, it has been checked, apparently by the arrival of deer.
The cedar belt on the lower edge of the bald is composed of conical trees, rather widely spaced, of an estimated age of 200-300 years, and in that respect unlike the commonly overmature cedar timber line in the Western Ruahine. The beech forest below is for some distance largely composed of mountain beech with an even canopy and appears to be a succession, presumably following fire, towards the red-beech forest of adjacent slopes.
On the bald the snowgrass cover is close, and as there is no seed source within at least a mile it is conceivable that tussock antedates the Tahuarongotea fire. Some Chionochloa conspicua is associated with it on its extreme margin; otherwise it is a practically pure stand.
The scrub is mainly but not entirely of a subalpine character. Leatherwood species are absent, though Olearia colensoi occurs rarely in forest in the vicinity. On the other hand manuka is frequent, unexpectedly at this altitude and 3-4 miles from the nearest known seed source. Coprosma rugosa, a characteristic divaricating shrub on river flats is another unusual component, and Olearia nummularifolia is recorded 10 miles south of its previously known limit
There is thus evidence that the Piringa bald originated mainly or entirely from an extensive pre-European fire spreading across the Waikamaka valley from the Mokai Patea plateau.
From the apparent age of the cedar it would have been at least 200 years ago, and from the depth of charcoal, not only on the slope, where it would have been partly buried by soil creep, but also on level ground on the spur of the Mokai Patea, the fire would have been appreciably earlier than this.
The mountain beech appears to have occupied the crest of the ridge more recently, perhaps 100 years ago, and the area of beech below the cedar appears to be of comparable age.
Daphne Burn. In the south (Map. 4) where the Daphne Ridge burn has run into the Oroua valley it has largely obliterated the Dacrydium-mountain beech ecotone. On its margins there are traces of the characteristic Dacrydium

40 Transactions – BOTANY VOL. 3
biforme in tussock but on the long spur SW from Taumataomekura it is cedar regeneration that is conspicuous in leatherwood scrub, with some scattered clumps of mountain-beech regeneration and unaffected red-beech forest below.
Otorere Beech Outlier. An area on the boundary of the Northern Ruahine in which mountain beech appears anomalously on a red tussock-cedar ecotone may conveniently be referred to here. The head of the Otorere Stream is a wide basin descending from the tussock plateau in the north into what was, from the patterns on either side of it, cedar forest. This has been replaced by a wedge of regeneration, much of it large Phyllocladus alpinus, apparently the result of a pre-European fire, the lowest point reaching the red-beech forest that occupies the centre of the valley. Although cedar is dominant and mountain beech absent over a considerable area on either side (as far as the Potae scarp on the one hand and Aorangi on the other), there are several stands of pure mountain beech near the upper timber line. Although cedar regeneration has been observed in a number of localities after fire, in this case mountain beech has come in, apparently after fire and apparently replacing former cedar forest.
WESTERN: SUBALPINE SCRUB
Scrub is generally absent from the northern part of the area. Leatherwood species are almost completely absent and in any case there are few points that rise above the timber line. One exception is Titapu (4,415ft), on a broad-topped ridge where an outlier of large Olearia colensoi has replaced former forest apparently Dacrydium biforme, to judge from the stumps still standing (1957) amongst leatherwood.
On the western slopes below Titapu Trig two shelves show up on aerial photographs which, from distant ground observation, appeared to be occupied by pure stands of leatherwood, probably on limestone remnants bracketed on to the greywacke ridge.
Across the Mangatera valley on the slopes of Trig T (4,715ft) large bushes of Olearia and partly moribund Dacrydium are scattered through snowgrass meadow almost to the summit.
On the Mokai Patea large bushes of Olearia present a similar pattern near Colenso Trig (4,613ft) and these share a curious feature with the large bushes near Titapu Trig, their bases having been recently (ca. 1956) recessed by browsing to a height of about two feet. On Titapu the leaves and terminals still lie untouched on the ground, with the twigs severed obliquely – presumably the work of hares. This has recently been seen also at other places on the range. Browsing of terminal twigs of Olearia oy deer, however, so extensive in the Tararua Range, has been noted only on an insignificant scale on the Ngamoko Range.
This pattern of dead or moribund Dacrydium biforme and large bushes of Olearia (with no young plants), scattered through snowgrass meadow at several points towards the northern limit of leatherwood and on a small scale on the Mokai Patea, suggests a general replacement of subalpine scrub-forest by snowgrass meadow.
In the southern half of the range leatherwood scrub is better developed, tending to form an almost impenetrable belt above the timber line, and where this is lowered, as at the head of the Oroua Valley and again at the southern extremity of the Ngamoko Range, may extend over a vertical range approaching 1,000ft. Towards this southern boundary characteristics of the Southern Rakhine begin to appear. Senecio elaeagnifolius becomes prominent towards the lower edge of the scrub belt and a Dracophyllum distinguished by its large juvenile leaves (noted under D. longifolium as Oliver’s var. septentrionale) makes its appearance.

TRANS. ROYAL SOCIETY OF N Z., BOTANY. VOL. 3, No. 3 PLATE 1
CENTRAL. WAIPAWA SADDLE. Contrast between the snow tussock slops to the Waikamaka on the west and the head screes of the Waipawa and Middle Creek on the east. The tussock triangle points to 3 Johns (extreme right).
(Photo: Crown Copyright, Land & Survey Dept.)



PLATES 4A AND 4B TRANS. ROYAL SOCIETY OF N.Z., BOTANY, VOL. 3, No. 3.
CENTRAL. Remnants of timber line (c. 4,700ft) on western face of main divide at the head of the Maropea River.
CENTRAL. BUTTERCUP HOLLOW. (4,400ft). Aged mountain beech with no sign of regeneration, the floor a closed community of co-dominant snow grass and Chionochloa conspicua.

TRANS. ROYAL SOCIETY OF N.Z., BOTANY, VOL. 3, No. 3 PLATES 5A AND 5B
NORTHERN. APIAS “BONEYARD”. Mainly Dacrydium biforme with numerous stumps in situ on weathered pumice subsoil, c. 3in below Taupo ash horizon. Present vegetation red tussock with mountain beech on better drained sites.
NORTHERN. Mountain beech hedge across red tussock plateau from near Aranga Hut looking N.W. across valley towards No Man’s Land. Celmisia spectabilis patch on left side.

PLATE 6A AND 6B TRANS. ROYAL SOCIETY OF N.Z., BOTANY, VOL. 3, No. 3
NORTHERN. NO MAN’S BOG. Face of Pit 11 showing Taupo horizon (point of knife) at 11in depth, Waimihia ash (tip of trowel) at 24-27in, with the latter rising over Dacrydium biforme lob (N.Z. 289 in 14C series) with remnants of grass plastered over it.
[Graph]
NORTHERN. NO MAN’S BOG. Positions of 14C dated timber with reference to surface.

TRANS. ROYAL SOCIETY OF N.Z., BOTANY, VOL. 3, No. 3 PLATES 7A AND 7B
WESTERN. AORANGI. Dense cedar-Dacrydium forest on limestone cap of Aorangi, also across the head of the Waipato Stream on an unnamed limestone block (right margin).
(Photo: Crown Copyright, Lands and Survey Dept.)
SOUTHERN. TAMAKI STREAM. Defoliated Weinmannia racemosa on hillside, dead Fuchsia, Aristotelia, and Neopanax on stream banks, rimu unaffected. May 1955.

No. 3 ELDER – Vegetation of the Ruahine Range 41
WESTERN: TUSSOCK AND FELL FIELD
These formations are practically confined to the Whanahuia and Ngamoko Ranges, whose easy rolling slopes carry extensive snowgrass meadows with the same species composition as the tussock of the central area apart from the appearance, in addition to Celmisia incana, of a form of C. discolor distinguished by its highly viscid upper leaf surface. C. hieracifolia has not been seen in the range and the reference to it m the Flora appears to refer to this species (A. P. Druce, pers. comm.).
Fell field is not extensive, carrying much the same assemblage of species as in the Central area, but the broad-leaved form of C. graminifolia growing closely appressed to the ground becomes relatively common.
WESTERN: BOG
There are patches of peat bog towards either end of the area, one near Titapu Trig in the north, the other on the Whanahuia Range in the south, both at altitudes of over 4,000ft, and both dominated by red tussock. Though neither is large they are of significance in that their species composition shows relationships with the larger bogs of the Mokai Patea and Northern Ruahine in spite of the distances between them.
The Titapu bog, as might be expected, shows affinities with the northern bogs though it has not the typical Sphagnum tarns (List 9).
The Whanahuia bog has some small tarns and shares with the No Man’s (NE) bog large quantities of preserved timber, also identified as Dacrydium of the biforme group, but unlike No Man’s, timber is thickest in the upper rather than the lower half of the deposit.
SOUTHERN RUAHINE (Map 5)
On either side of the Manawatu Gorge the same forest and scrub patterns recur in the Tararua and Ruahine Ranges, and can be visualized as mirror images of each other. Podocarp-hardwood forest occupies the lower slopes on both sides of the gorge, and as the land rises kamahi forest becomes dominant above this, merging into leatherwood scrub associated with more or less Dacrydium biforme. At a distance of 15-20 miles from the gorge in either direction red beech begins to replace kamahi, and still farther away most of the leatherwood belt is also replaced by beech forest, silver beech to the south, mountain beech to the north.
The meteorological factors appear adequate to account for the close correspondence of these patterns. This cloudy and windswept part of the range, the Southern Ruahine, is also a distinct topographical unit, separated from the rest of the range by the Pohangina Gorge and the low (c. 2,500ft) Apiti-Norsewood Saddle. It is for the most part a long plateau tilted away from an abrupt scarp on the eastern side to a less defined scarp on the west, and barely exceeds 4,000ft at its highest point, Takapari.
Ecologically it is a well defined natural area, the vegetation falling into three main altitudinal belts, podocarp-hardwood forest, kamahi forest, and leatherwood scrub. There are also vestiges of two other formations, cedar-Dacrydium forest and snowgrass tussock.

42 Transactions – BOTANY VOL. 3
These belts do not precisely match the equivalent belts occurring on the rest of the range. The most obvious difference is that a number of characteristic species show a definite drop, of the order of 500ft, in their upper limit. Rimu, Podocarpus hallii, cedar, Dacrydium biforme, and Senecio elaeagnifolius, with Coprosma foetidissima and Chionochloa conspicua are all in this category. Other physiognomic species show no appreciable change, but on the other hand one important dominant, kamahi, reaches a considerably higher altitude than farther north. This redistribution of species is presumably due to climatic influences, of which light, humidity, and wind are the most suggestive.
The vegetation is accordingly described in five altitudinal belts, three major and two minor, the latter being included on account of their presumed historical significance. As animal damage has changed and is changing the vegetation so fast, some point of time must be chosen at which it can be described. The difficulty is that the period 1955-1960, when most of the detailed investigation was made, covers too many changes; accordingly species distribution is based as far as possible on observations made before 1945, when animal damage was still sporadic.
SOUTHERN: RIMU-RATA FOREST (1,200-2,300ft)
Large emergent trees of these two species give the lower forest its character but a considerable variety of other species is associated with them. Some of these only appear along the lower fringes and are representatives of lowland forest proper which survives in the Totara Reserve (Greenwood, 1949). Some species have the vertical range of rimu and rata or may exceed it, but most have dropped out at an intermediate level, so that the associated forest species fall into three groups-below 1,200ft, below 2,000ft, and up to 2,300ft or occasionally higher (List 14).
In comparison of the 1,200ft group with the Totara Reserve forest, Eugenia maire has not been recorded and Laurelia novae-zelandiae recorded only in one small swampy hollow.
The most interesting common species is black beech (Nothofagus solandri), which occupies the driest sites on the Pohangina River. It occurs on sandstone blocks at the mouth of Te Ekoau Stream and also forms pure stands on spurs farther upstream and in the adjacent Makawakawa valley. The uniform canopy of these beech stands contasts [contrasts] sharply with that of the neighbouring forest, and particularly so in the absence of other species of beech from the whole area. I understand that these are vestiges of a much more continuous black-beech forest which formerly capped the long ridge between the Oroua and Pohangina valleys (B. E. Sixtus, pers. comm.).
In the middle and largest group (below 2,000ft) tawa, though common and in suitable locations plentiful, has a high proportion of saplings and rickers which seldom reach the canopy – recent invader but by no means an aggressive one.
Abundance of lianes and epiphytes is characteristic of this belt, particularly of supplejack (Ripogonum scandens), whose stems form thickets, at their worst impenetrable to humans, over considerable areas, while its leafy upper portion typically sprawls over the lower canopy between the tallest emergents, supplying for instance a considerable proportion of the cover in aerial photographs.
SOUTHERN: KAMAHI FOREST (below 3,000ft)
The drastic changes brought about in this forest by the arrival of opossums, with some assistance from goats and latterly deer, make a general description almost impossible. Prior to 1950 kamahi was the dominant canopy tree from the limit of rimu up to about 3,000ft; associated with Podocarpus ferrugineus in the

No. 3 ELDER – Vegetation of the Ruahine Range 43
lower part of its range and P. hallii in the upper. Undergrowth both of shrubs and ferns is dense, but recent lists (List 15) show a high proportion of unpalatables and raise the suspicion that animals may have modified the vegetation over a longer period than was earlier appreciated.
A few species of minor importance, such as Libertia pulchella and the semiepiphytic Enargea parviflora, have a conspicuously narrow altitudinal range and are characteristic of this belt. In moist sheltered hollows Todea superba may escape browsing, and one curious fern, Blechnum nigrum, found only in this part of the Ruahines, is apparently confined to the darkest sites in a narrow belt at about 2.500ft.
SOUTHERN: LIBOCEDRUS-DACRYDIUM FOREST
This is no more than the ghost of a formation at the present day, but it may approach dominance at about the 3,000ft contour often enough to afford a comparison with the same community further north, of which it is assumed to be a relic. Elements of it can be traced upwards almost to the crest of the range (c. 3,600ft) and downwards on spurs and knolls as far as the boundary of podocarp-rata forest.
SOUTHERN: LEATHERWOOD SCRUB
With the whole of the crest (which forms a wide plateau in the northern half) above the timber line, perhaps as much as 20% of the Southern Ruahine is occupied by leatherwood scrub. The two dominants, Senecio elaeagnifolius and Olearia colensoi are more distinctly separated than elsewhere m the Ruahines or Tararuas into a lower Senecio-dominant and upper Olearia-dominant zone. Although most of the other species of the subalpine scrub association are present they occur as isolated plants, hardly breaking the uniformity of the canopy. Such characteristic species as Coprosma pseudocuneata, Pittosporum rigidum, Olearia ilicifolia, Neopanax colensoi, N. simplex var. sinclairii, and Aristotelia fruticosa var. are rare. Olearia arborescens is still frequent, although it ranks high as a palatable species. A sizable colony of Phormium tenax in flower in January just below the main leatherwood zone at 3,100ft is a strange feature on one spur to the east of Matinginui.
SOUTHERN: SNOWGRASS (Chionochloa pallens)
While the limited occurrence of snowgrass hardly merits a separate category, it is of some ecological significance and has accordingly been distinguished. It occurs on Takapari, at the 4,000ft mark, interspersed with Olearia scrub, forming the pattern common in other parts of the range. The two formations appear to coexist, the leatherwood growing outwards and downwards (as is its habit) into tussock and killing it by shading it, probably assisted by snowdrifts in winter, while on the uphill side of the bush the exposed stems are laid open, in their tum, to invasion by snowgrass; the alternate strips of snowgrass and leatherwood apparently succeed each other down slope in a series of waves.
Ten miles farther south, on either side of Maharahara Trig, strips of tussock dominate exposed ridges at about 3,200ft, where they appear to be replacing leatherwood. North of this two or three grassed basins one or two acres in area show up on aerial photographs in leatherwood on the broad plateau between

44 Transactions – BOTANY VOL. 3
Maharahara and Takapari. Access to these is difficult, and their status is still uncertain. Their pattern suggests tussock bog, and in the Northern Ruahine such boggy basins are occupied by red tussock (Chionochloa rubra). A recent Forest Service aerial reconnaissance took especial notice of these southern tussock basins and considers that there is a strong likelihood of their being composed of red tussock. If so they form a link between the main area of red tussock in the central North Island and the outliers found on Kaiparoro and Taramea in the Tararua Range (Druce, 1957).
COMMENTARY
What marks the Ruahine Range out from all other North Island ranges is the amount and variety of visible changes that have taken or are taking place in the plant cover.
The changes can be put into some sort of perspective by the evidence preserved in peat bogs, several of which have been sampled for pollen profiles, but on the other hand the processes of change taking place in the living vegetation have been so complicated by the arrival of deer and other browsing animals that until closer study is possible interpretation must remain tentative.
PEAT PROFILES
The information contained in the peat bogs is fundamental. Various ash showers are present in narrow bands, the latest of which are identifiable, and these are available for dating. The earlier have not yet been accurately determined, but the deeper peats may be taken to cover at least half and probably most of the post-glacial period (Moar, 1961).
Pollen analyses for the four main Ruahine bogs (Reporoa, No Man’s Land, Mokai Patea, and Whanahuia) agree in showing a general change from podocarp to beech over a period of several thousand years, but the detail differs from bog to bog so that no very close comparison is yet possible between the histories of different parts of the range. The bogs lie at elevations between 3,800ft and 4,700ft – that is above or level with the present timber lines in their neighbourhood, so that the historical problems are primarily those of the relations between mountain beech, Libocedrus bidwillii, and Dacrydium biforme, to a secondary degree between these and red beech, and to a minor degree with silver beech.
The pollen of the last is distinct and its sporadic occurrence over a long period in the western bogs is a valuable piece of evidence that at no stage has it been a serious competitor to mountain beech, but as none of the other beech pollens are distinguishable from one another no comparison of the ratios between red and mountain beech is possible.
My other difficulty is that the frequency of a pollen bears no certain relationship to the frequency or propinquity of the species. Thus Dacrydium biforme timber shows clearly that it has been abundant for thousands of years on the No Man’s and Whanahuia bogs, yet pollens of its group are far less conspicuous in a profile than those of one lowland species, rimu (D. cupressinum), trees of which could only have been brought into the vicinity of the bogs by a climatic upheaval which would have disrupted the pollen profiles. The extreme case is cedar, whose history is of the highest importance as it takes the place of mountain beech near all the western bogs, but whose pollen is absent except for traces recorded in the Reporoa peats.

No. […] ELDER – Vegetation of the Ruahine Range 45
LIBOCEDRUS/DACRYDIUM BIFORME RETREAT
The longest-lived trees in the upper forest are the gymnosperms, and their condition gives the clearest indication of change. Libocedrus and Dacrydium communities show a consistent pattern of deterioration which is most advanced in the south, in a sequence which appears to cover several generations and to follow to some extent a sunshine gradient northwards. Both are favoured by limestone in the NW and both show some pioneering vigour under certain circumstances, notably after exposure of the ground by fire or by opening of the canopy.
LOWLAND PODOCARP REMNANTS
Most lowland forest has been almost completely destroyed, so that little can be deduced from the state of the podocarps in the fringes that remain. The persistence of totara-matai forest on the Rangitikei River below Aorangi suggests a connection with the presence of limestone, and a combination of deep gravels and low rainfall between the range and the Hawke’s Bay plains has probably had a similar effect in preserving the former Seventy Mile Bush against the encroachment of hardwoods. Both totara in grassland and rimu on burns are capable of abundant regeneration although, in general, both species, together with matai, are failing to replace themselves under forest conditions.
MOUNTAIN BEECH DETERIORATION
The main and so far unsolved problem of the Ruahine Range is the general deterioration of mountain-beech forest except towards the lower end of its range, where it is healthy and aggressive. The retreat of its timber line 130 miles north on the Maungawaro Range suggests a general change of environment unfavourable to the species, but this is not borne out by its good condition in the volcanic areas. In the Ruahine Range itself the relationship between mountain and other beech species is contradictory. There is sporadic evidence of a corresponding retreat of red beech and of its replacement by mountain beech, but where silver and mountain beech meet on the western side of the range the two species coexist in all-aged groups at the present day, with silver beech as a minor component. Peat profiles show that this has been its status, though intermittently, over a period of several thousand years. It is difficult to conceive of a climatic fluctuation of any significance that would not affect the balance between these two species. It is possible that silver beech is in retreat elsewhere; but the evidence is contradictory and in any case is masked by the longer life span of silver beech which enables it to ride over short-term fluctuations affecting mountain beech.
The comparatively short life span of mountain beech (c. 150 years), its rapid decay, and the impracticability of distinguishing its pollen from those of the rest of the N. fusca group tend to limit evidence to events of the past 200 years; in fact, as with gymnosperms, one tends to accept a time scale which is set by the life of an individual of the species. For this reason investigation of the vulnerability of mountain beech emphasizes recent catastrophe of one form or another with little sign of recovery. The only evidence of a longer period of decline could come from the persistence of the lowered timber lines on the Mokai Patea plateau if these were found to date from an earlier period of Maori occupation.

46 Transactions – BOTANY VOL. 3
CATASTROPHES
Most of the conspicuous discontinuities that can be recognised in the present vegetation are in the nature of catastrophes which do not link up with one another to suggest steps in a general process but force consideration of the importance of fortuitous disasters, sometimes with long-lasting effects, as normal hazards of plant communities, particularly as they approach their limits.
Earthquake
No damage has been seen due to the Hawke’s Bay earthquake (1931), though some could have been expected considering the extensive slip and rock-fall damage that followed the Murchison quake (1929).
In the upper Makaroro the frequency of steep rubble slopes in second growth in the vicinity of a sharply marked fault trace has suggested a connection, but this has not been confirmed. As further north in the Kaweka Range (Makino Valley), earthquake damage along the East Coast fault system remains no more than a probability.
Cloudbursts
On at least two occasions since the Hawke’s Bay earthquake, damage caused by heavy local rainfalls has been far more obvious in river valleys than any earthquake damage. Neither of these flash floods attracted attention outside the range, but the more recent of them (September, 1954) destroyed numbers of terraces in three valley heads which are estimated, from the size of the timber growing on them, to be 200-300 years old.
Fire
Traces of pre-European fire are definite on the Mokai Patea plateau and fairly certain in two areas of the Northern Ruahine. The forest patterns are still clearly modified, and the subsurface charcoal on the Mokai Patea may be taken as likely to date within the period of Maori occupation on the analogy of the recently investigated South Island charcoals (Molloy et al.).
One of the earliest of the European fires, the Daphne burn of the 1880s, similarly appears to have initiated a non-reversible change to snowgrass or leatherwood scrub in place of the Conner mountain-beech timber line, with, however, some stimulus to the regeneration of Libocedrus.
Gale
There is only localized evidence that can be certainly put clown to exceptional winds. Between the Makaroro and Waipawa rivers knolls and exposed spurs in red-beech forest are conspicuous from their close wind roof of mountain beech. Under this lie large red-beech logs, but beheaded trees, some of them of large size, are still alive, their branches merging with the smooth canopy. Some more widespread damage, particularly in mountain-beech forest, has features resembling some of the less obvious patterns of damage in the Tararua range as a result of the 1936 gale. This gale incidentally was of little significance in the Ruahine Range, except perhaps in the Pohangina valley, where it brought down trees in the Totara Reserve and may account for some of the wind throw in Weinmannia forest on the middle slopes of the range south of the Opawe Stream.

No. 3 ELDER – Vegetation of the Ruahine Range 47
Snow
Damage due to the lodgment of heavy falls of snow had not been particularly noted until the winter of 1961, at any rate as distinct from gale damage. This fall was unusually widespread and particularly affected red-beech forest, causing heavy damage in several localities on the western side of the range.
Animals
Animal damage has been most spectacular in the almost complete defoliation of the canopy by opossums over large areas of Weinmannia-dominant forest in the Southern Ruahine (Plate 7B).
Over the whole range, ground-browsing animals, mainly deer, have had the greatest effect on beech forest, mainly on the seedlings of mountain beech, but have not yet turned seriously to the leatherwood scrub belt. Ground- and canopy-browsing animals each improve the habitat for the other, but protection measures came early enough in the history of animal invasion to check an uncontrolled population build-up over most of the range, though at either extremity opossums (Southern), deer (Northern), and perhaps goats (in the Wakarara Range) had passed their greatest density before planned control started (Appendix 1).
ACKNOWLEDGMENTS
Messrs A. P. Druce and R. M. Greenwood have collaborated with me and we have shared much of the fieldwork on which this paper is based. In unravelling the history of the complicated changes in the vegetation I have been much indebted to persons with long knowledge of certain areas, in particular to Messrs E. S. Klein, F. and K. Severinsen, B. E. Sixtus, R. A. L. Batley, E. H. Beamish, the late J. B. Campbell, and the late L. Masters.
From 1938 onwards I have been indebted to Government hunters, originally of the Department of Internal Affairs and latterly of the Forest Service, for up-to-date information, particularly about the state of various remote areas, and since 1954 I have been employed by the N.Z. Forest Service on work of which this paper is one aspect. My thanks are due to the department and to those of its officers who are concerned with problems relevant to this paper; to the Botany Division and the Geological Survey of D.S.I.R, and to the Meteorological Office for help in particular instances.
APPENDIX 1
INTRODUCED ANIMALS: ARRIVAL AND SPREAD
Rabbits
The first record of wild animals having appeared in the range is contained in a letter dated January) 1900 from Mr R. T. Batley (Moawhango) to the Hawke’s Bay Rabbit Board which reports the appearance of rabbits on the Mokai Patea plateau, and states that these were the first to have reached the western side, having apparently filtered across the range by way of a new track cut through the bush on the Hawke’s Bay side. This was presumably the pack track to Shut Eye Shack, near the head of the Waipawa River. This hut, together with “Hut Ruin” at the head of the Makaroro and No Man’s Hut, appear to have been in a chain of huts for a rabbit board patrol along the northern part of the range. There were also rabbit board huts at the head of the Pohangina and on the Mokai Patea plateau.

48 Transactions – BOTANY VOL. 3
During the 1914-18 war an upsurge of rabbits over the Inland Patea affected both Mangaohane and Otupae seriously. On Otupae it took an intensive three-year campaign to suppress them and a further four years before they were completely eliminated (J. B. Campbell, pers. comm.).
Within the range small colonies of rabbits have been seen intermittently over the past 25 years, mainly on the NE plateau and on Weka Flat in the upper Kawhatau valley. My most recent records are No Man’s (October 1956) and Weka Flat (January 1961), and rabbits have also been glimpsed at the head of the Mokai Patea plateau just below Rongotea trig (January 1961).
Red Deer (Cervus elaphus)
The two earliest red deer liberations in the North Island were in the Wairarapa (3) in 1862, and at Matapiro (11) in 1883. The latter, only six miles east of the range on the northern bank of the Ngaruroro, is clearly enough the main source of the Ruahine herds, though it must be noted that as early as 1867 or 1868 a stray stag which could only have come from the Wairarapa herd had appeared on Tutira, well to the north of the Ruahines (Guthrie-Smith, p. 336).
By 1890 a few deer had reached the spurs of the range behind Kereru, the nearest point to the Matapiro liberation (L. Masters), while a cast antler on Deer Horn Hill (1900) farther south was equidistant from both liberations (K. Severinsen, pers. comm.).
On Whanawhana, immediately north of the range and across the Ngaruroro, the first deer were not seen until 1906, were becoming a menace by 1910, and reached their peak of population in the late 1920s; numbers started to decline in the mid-1930s (E. H. Beamish, pers. comm.).
On the range itself deer were reported above the timber line on No Man’s Land by 1910, and tracks were seen as far south as Tiraha at the head of the Tukituki in that or the following year. Deer were also reported on the western side of the range (Mokai Patea, Mangaohane) from 1910 onwards (L. Masters, F. Severinsen, T. V. Morrin).
There were three small liberations on the western side about this time, in the Kauwhatau (5) in 1908, in the Oroua (4) in 1914 and south of the Pohangina at Delaware (4) in 1922. These can have been little in advance of the first spread of animals from the northern end of the range.
By the mid-1920s there was a dense population on the northern plateau; the forest floor was bared and roots were exposed (L. Masters). Between 1933 and 1935 there was evidence of a fairly rapid southward shift accompanied by the development of a strong track system, particularly towards the head of the Waipawa River. By 1935 there was sign of deer along the whole line of tops from the Ngamoko Range to Kereru, with a steady increase as one proceeded from south to north. In this year exposure of the forest floor by trampling became particularly noticeable in the head of the Waipawa, and from 1939 onwards new slips in both the Waipawa and the Makaroro Rivers have been frequent.
Kean and Newcombe’s report (1937) on a crossing of the range early in that year shows clearly that deer had then reached a more advanced stage of occupation in the extensive western valleys than on the divide itself and the eastern slopes. This report repays close study at the present day as it gives detail about browsing patterns and preferences from evidence which is no longer available.

No. 3 ELDER – Vegetation of the Ruahine Range 49
Internal Affairs shooters first visited the range in 1938 and found that most of the deer were then to the north of the Waipawa River.
By 1940 there was a heavy concentration of deer in the upper Pohangina valley, but only a few strays south of this. From 1945 onwards to 1962 there have been consistent reports of a build-up of deer throughout the Southern Ruahine.
Sika Deer (Cervus sika)
These are commonly called Japanese deer though specifically distinct. Their habits differ from those of red deer and in consequence they have different effects on the vegetation and present more difficult problems of control. They have only recently (1957) made their appearance in one limited area of the range (Makirikiri headwaters). This was a continuation of their previous rapid spread from the north, down the eastern flank of the Kaweka Range, but apart from a stray stag shot near Gwavas in 1961 (T. Cookson, pers. comm.) there has been no indication of their further penetration. Two stags were heard at the Makirikiri in the 1961 roar but the indications are that this remains an isolated small colony whose effect to date has been negligible.
Goats
Goats occur at either end of the range and, until recent hunting operations, were spreading inwards at a rather slow rate.
In the Southern Ruahine goats were first seen in the head of the Raparapawai as early as 1919 and were then thought to have crossed the range from the vicinity of the Opawe (S. Klein, pers. comm.), though the earliest introduction recorded there is 1925. From here they spread north and south either way from the Raparapawai at a fairly slow rate – it was not until 1955 that they had reached the southern extremity of forest at Wharite; to the north they scarcely put in an appearance in the Tamaki valley on the eastern side, while on the western side they have appeared in the upper Pohangina valley only since 1959.
In the northern Ruahine goats established on the Wakarara Range had spread across Duff’s Flat on to the main range by 1954 and were working south beyond the Makaroro River by 1958.
Opossums
Information about the liberation and spread of opossums is very fragmentary and almost certainly confused by private liberations during the period of licensed trapping. They appear to have first become established at either end of the range and were first known to have become established on the eastern side (Oruakeretaki) in 1923 (S. Klein). They were in the Wakarara area towards the northern end before 1937 and in that year made their appearance towards the centre of the range at Ashley Clinton (K. Severinsen, pers. comm.)
The subsequent pattern of their spread shows a remarkable uniformity over a good part of the range. High populations built up on the fringes of the forest, and it was at least five years in the Tukituki and much longer elsewhere before evidence of their spread into the higher beech and Weinmannia forest became apparent. Between 1945 and 1955 the upward movement of opossums was noted from several parts of the range. In the winter of 1955, there is a suggestion of collapse of the opossum population above the Pohangina valley (SW Ruahine).

50 Transactions – BOTANY VOL. 3
The following winter there was a remarkable incursion of opossums into huts on or near the timber line along the northern half of the range from No Man’s to the Waikamaka, and accompanying this repeated reports of opossum tracks and signs at the timber line, particularly in snow.
Rats and Mice
Rats are commonly established near huts, but over a period of nearly 30 years I have seen little sign of them in the bush or round temporary camps. The first observations of mice were made in the Makaroro area in the winter and spring of 1955. They were repeatedly remarked on all through the forest in October and November of that year, one informant stating that drowned mice were frequently seen in the river and that a bush-line camp was infested with them (W. Romanes, pers. comm.). In the following new year mice were abundant in the Mokai Patea tussock (B. G. Hamlin, pers. comm.) and there was a sizable colony at No Man’s Hut. Over this period a similar eruption occurred in the Kaweka and Kaimanawa Ranges, and a connection between this and the mast fall of the previous autumn may be postulated. Numerous mouse plagues, some associated with mast years, are recorded from the South Island, but only one previously from the North Island, at Rangataua and apparently following a mast year in 1949.
Pigs
Fifty years ago pigs are said to have been abundant in some of the eastern valleys of the Southern Ruahine but to have almost completely disappeared since. Over the past 25 years a scattered and apparently fluctuating population has been observed on the lower slopes throughout the range.
Mustelids
Three stoats were observed playing together in the Herrick’s hut area in November 1955 (W. Romanes). To my knowledge this is the sole record of any species.
INTRODUCED ANIMALS: EFFECTS
Deer
As early as 1932 mountain-beech forest was already a shell on the slopes of the NE plateau, which was the area of the densest deer population. Nearly all undergrowth had then been browsed out in some basins, even the beech seedlings.
As deer population increased southwards along the range the pattern repeated itself in the forest, and by 1937 damage to alpine scrub was apparent in the head waters of the Waikamaka and Kawhatau Rivers (Kean and Newcombe). Some of it, such as the browsing of Olearia colensoi, is surprising at such an early date, while the barking of Pittosporum rigidum is quite unusual, being in fact generally regarded as browse resistant. An observation of importance at that date is that Blechnum capense on valley walls was then rapidly disappearing. This fern typically flourishes on steep faces of loose soil which are vulnerable to slipping. There is little evidence today that it was ever an important species.
In the Makaroro area recent investigation shows a marked gap in beech regeneration between 1930 and 1940 which coincides with the apparent peak of deer population (Widdowson, Ellis).

No. 3 ELDER – Vegetation of the Ruahine Range 51
By 1940 the eastern valleys, Waipawa and Makaroro in particular, were developing slips and screes on steep forested slopes. These had been initiated by trampling, which was destroying the mat of mosses and fine rootlets and allowing the rock beneath to break up. The stabilization and revegetation of slip surfaces has been slow, so that it appears that a much reduced deer population can delay healing almost indefinitely. Below a certain level, however, recovery can be rapid.
Deer were rarely seen in the scrub forest of the Southern Ruahine before 1945, but more recently the defoliation of the canopy Weinmannia by opossums has allowed a luxuriant understorey to come away, much of it palatable to deer, and a steady build-up of deer in that area is still being reported.
Hares
These are not as common as they are on the Kaweka Range, where they are abundant, and their influence is interesting rather than serious. Browsing of Aciphylla was attributed to them (Kean and Newcombe), and in the last five years or so the undercutting of bushes of Olearia colensoi to a height of 2ft has been noticed in a number of localities. This was almost certainly the work of hares.
Goats
Goats are more localized than deer and they browse more intensively, showing a strong preference for open spaces exposed to the sun, which they hammer unmercifully. Their fondness for Geniostoma is marked, but their taste is so catholic that an evaluation of species browsed would be pointless.
Their spread throughout the Southern Ruahine, which was perhaps encouraged by their disturbance in the early stages of the extermination campaign, has probably contributed in some degree to the rapid spread of opossums a little later.
Opossums
The spectacular canopy defoliation over the Southern Ruahine developed over so short a period that it has only been sporadically observed, and unfortunately the changes were gathering momentum during the war years when practically all hunting and botanical work was halted, so that many of the processes of change are obscure. To add to the complexity of the situation abrupt changes in the animal population since have introduced new factors, not only the collapse of an opossum population but also the resumption and intensification of Forest Service hunting and the initiation of 1080 poisoning trials. With the possible exception of the podocarp-hardwood forests along the margin of the range the present plant cover of the Southern Ruahine appears to be in a highly unstable state. Observations become out of date soon after they are made, and its future development is a matter for surmise.
Fortunately Messrs Druce and Greenwood had covered most of the Southern Ruahine prior to World War II and their botanical reconnaissance gives an invaluable general picture of the distribution and frequency of species before the animal invasion.
Aerial photography in October 1946 came at a useful stage, when the forest canopy was still substantially intact, at least in the Weinmannia belt, though the texture on the lower slopes suggested extensive crown fires to the Forest Research

52 Transactions – BOTANY VOL. 3
Institute interpreters. It now seems probable that defoliation of rata could have given this impression. (It is, however, on open question how far the death of rata can be attributed to opossums; its death on Kapiti (1907) and its sudden death over a short period at Waikanae (1936, D. Port, pers. comm.) appear to have other causes.) In the Southern Ruahine the death of individual trees of this and other species was noted as early as 1948 (Pracey), and by 1952 defoliation of rata and Weinmannia was on a large enough scale to be visible from the plains (Klein). By 1955 this was general. Investigation showed considerable changes in forest composition with no recognizable cause except opossum browsing, of different intensities, over a wide range of species that showed varying rates of resistance (List 16).
Patterns of opossum browsing can only receive brief mention here so far as they affect vegetation. Opossums show a strong preference for leaves fully exposed to light, probably because leaves in the canopy are either more nutritious or more closely packed and accessible. Only two tree species, Beilschmiedia tawa and Myrsine salicifolia, have been observed to be browsed significantly while in the sub-canopy. Individual trees, particularly those of rata, can be observed that have been killed or almost completely defoliated while their neighbours are almost untouched. This is consistent with the comment from residents of the Pohangina valley that concentrations of opossums appear on rata as soon as leaf growth starts. It seems that browsing, by stimulating the production of fresh shoots, would prolong concentration on an individual tree to the point of its exhaustion.
Pseudowintera colorata is so definitely unpalatable to all animals that its abundance in the undergrowth of beech forest is a recognized effect of former browsing. Its barking by opossums, first noticed as early as 1948 (Pracey) on a major scale, has now reached serious proportions in the Southern Ruahine. (There is one recent record of it elsewhere in the NW Kaimanawa Ranges Mackenzie, pers. comm.) It is noticed that it accompanies a heavy local browsing pressure, suggesting a dietary deficiency of some sort. Browsing of Griselinia littoralis (hitherto considered completely unpalatable to opossums though not to deer) has been noted in the Central Ruahine though only in one locality, and may indicate some similar type of deficiency.
The occurrence of small stands of vigorous and almost completely unbrowsed Weinmannia in the middle of dead and defoliated forest cannot be satisfactorily explained. All have one common feature – the site, typically level or gently sloping, in a broad saddle or hollow, probably with a deep accumulation of humus and rather damp. Several such pockets have been examined in the southern Ruahine and at least one is known in the Tararua Range.
As far as can be judged at the moment (1962) the marginal forest of the Southern Ruahine is fairly stable, trapping having probably helped to keep the opossum population down. Above this the defoliation of Weinmannia has allowed a rush of palatable undergrowth to come away, too dense for goats but favourable for deer, which are tracking and occupying the uppermost leatherwood scrub belt though this is not yet much affected.
Two deer- and goat-proof enclosures have been sited in the Opawe area, one at 3,100ft on the forest-scrub boundary, the other at 2,000ft in former kamahi forest, with the aim of keeping a check on rapidly changing conditions.

No. 3 ELDER – Vegetation of the Ruahine Range 53
INTRODUCED ANIMALS: CONTROL
As the effect of browsing animals has become an important factor in any study of the vegetation an outline of the development of control operations is of some value.
Deer
Operations in the first place were directed against deer. Shooting by the Department of Internal Affairs began in July 1938, and in the first season some 5,000 were tallied in the Ruahines, mainly in the northern half. War stopped the programme and when shooting was resumed it was found that the deer population had decreased, at least in the northern half, which had been well eaten out. Since then shooting tallies, except for one or two minor increases, agree with other evidence of a general decline in numbers, most apparent in the north (P. Logan, pers. comm.).
By 1955 there was some evidence of the recovery of vegetation, but not over the whole area. It appeared that on the whole a lower deer population was sufficient to hold the vegetation in a depleted state and that control by existing methods had reached a stalemate.
In consequence the attempt at overall control was abandoned in 1958 in favour of a concentration of the available manpower on areas selected as being of most importance from the point of view of erosion, and these were intensively shot by hunters working methodically in teams over 10-11 months of the year. Considerable tallies have been taken and there is evidence of the recovery of vegetation, but the extent and rate of this requires closer study. Now that animals have become scarce in the highest priority catchments, the pressure of hunting tends to be transferred to adjacent buffer zones to prevent the infiltration of deer. The four or five years following the next beech-mast year (from present indications, 1963) will be vital for the maintenance of mountain-beech forest.
Goats
Goats were abundant and deer scarce in the Southern Ruahine at the end of the war, and between 1951 and 1956 goat hunting, sometimes with the use of dogs, was carried out. By 1958 goats were widely scattered in no great concentrations and recovery of vegetation was evident. Experience with teams using dogs has shown that the extermination of goats is practicable, and the arrest of their northward spread in the Southern Ruahine Range has been a first priority since 1958.
In the Northern Ruahine the arrest of their spread south of the Makaroro has been similarly the first priority and the Gold Creek colony was eliminated before proceeding to clear the rest of the area. This has not been completed as both goats and wild sheep still occur on privately owned land at the northern extremity of the range.
Wild Sheep and Pigs
Experience has shown that the methods used by goat-shooting teams are equally effective against both sheep and pigs, but in the Ruahines neither are of great importance except for sheep on private property along the Taruarau valley on the northern border, where an appreciable number are said to have drifted back from farm land in the drought of 1946 (L. Masters, pers. comm.).

54 Transactions – BOTANY VOL. 3
Opossums
An aerial poisoning trial using 1080 over 1,500 acres of podocarp-rata and Weinmannia forest at the head of the Opawe Stream on the western side of the southern Ruahine was carried out in February 1959. From check trapping a 74% kill was estimated and there has since been evidence of recovery of foliage. A higher kill, however, is considered necessary for effective control.
APPENDIX 2
VERNACULAR NAMES
In the text locally familiar names are used in the case of a few prominent species which are being frequently referred to. The following list will serve to identify them.
VERNACULAR NAME FAMILY SPECIFIC NAME
Podocarpaceae
Kahikatea Podocarpus dacrydiodes A. Rich.
Matai P. spicatus R.Br.
Miro P. Ferrugineus D. Don
Totara P. Totara D. Don
Rimu Dacrydium cupressinum Soland.
Cupressaceae
Cedar (kaikawaka) Libocedrus bidwillii Hook. f.
Myrtaceae
Manuka Leptospermum scoparium Forst.
Kanuka L. ericoides A. Rich.
Rata Metrosideros robusta A. Cunn [Cunningham].
Elaeocarpaceae
Wineberry Aristotelia serrata (J. R. et G. Forst.) W. R. B. Oliver
Cuconiaceae
Kamahi (tawhero) Weinmannia racemose Linn. f.
Papilionaceae
Kowhai Sophora tetraptera J. Mill
Fagaceae
Silver Beech Nothofagus menziesii (Hook. f.) Oerst.
Red Beech N. fusca Oerst
Black Beech N. solandri var. solandri (Hook. f.) Poole
Mountain Beech N. s. var. cliffortiodes (Hook. f.) Poole
Oleaceae
Maire (black) Gymnelaea cunninghamii (Hook. f) L. Johnson
Maire (white) G. lanceolata (Hook. f.) L. Johnson
Compositae
Leatherwood Olearia colensoi Hook. f
Senecio eleagnifolius Hook. f.
Raoulia spp.
Gramineae
Snowgrass Chionochloa pallens Zotov
Red tussock C. rubra Zotov
Smilacacae
Supplejack Ripogonum scandens

No. 3 ELDER – Vegetation of the Ruahine Range 55
PLANT LISTS
In the following lists I have indicated, where sufficient information is available, my estimate of the abundance of each species on the following basis:
5 = dominant or co-dominant
4 = abundant or plentiful
3 = common
2 = a minor but recognisable component of a community
1 = rare
loc = local
ep = epiphyte
Names, except for Monocotyledons, follow Allan’s Flora (1961).
LIST 1
ENDEMIC TO RUAHINE RANGE
(Hebe truncatula)
Epilobium gracilipes var. concinnum (NW Ruahine)
ENDEMIC TO INLAND PATEA AND RUAHINE
Myosotis eximia
Hebe colensoi var. colensoi
Hebe colensoi var. hillii
Epilobium pubens var. astonii (also Raukumara Range)
ONLY NORTH ISLAND LOCALITY
Senecio glaucophyllus subsp. discoideus
Geum leiospermum
(Gaultheria crassa)
Also a number of species listed by Druce from the Reporoa Bog, amongst which are:
Euphrasia disperma
Myosotis tenericaulis
Ranunculus depressa var. stewartiae
Ranunculus ternatifolius (Makirikiri)
LIST 2. GENERAL. FOREST below 2,000ft, MINOR SPECIES
CANOPY
Elaeocarpus dentatus
Weinmannia racemose
Gymnelaea cunninghamii
Gymnelaea lanceloata
SUB-CANOPY
Cyathea dealbata
Pittosporum eugenoides
Schefflera digitate
UNDERSHRUBS
Geniostoma ligustrifolium
Coprosma robusta
Helichrysum glomeratum
EPIPHYTES AND PARASITES
Asplenium falcatum
Elytranthe flavida
Earina mucronata
Earina autumnalis
LIST 3. GENERAL. FOREST SPECIES
(These lists are drawn up to give a general picture of the composition of the central Ruahine, but they apply with minor modifications to both northern and western areas (cf. Lists 8 and 12). To some extent they apply to the southern Ruahine, but the differences are greater (cf. Lists 14 and 15) ).
A. Upper Limit 2,000-3000ft B. Upper Limit 3,000-4,000ft
CANOPY
Podocarpus spicatus
Podocarpus ferrugineus
Dacrydium cupressinum
Podocarpus hallii
Libocredus bidwillii
Nothofagus fusca
SUB-CANOPY
Clematis paniculata (liane)
Clematis forsteri (liane)
Melicytus ramiflorus
Pittosporum tenuifolium
Aristotelia serrata
Carpodetus serratus
Neopanax arboreum
Pseudopanax crassifolium
Coprosma linariifolia
Coprosma australis
Cordyline banksii
Olearia rani
Fuchsia excorticata
Elaeocarpus hookerianus
Rubus cissoides (Liane)
Neopanax simplex
Neopanax simplex var. sinclairii
Griselinia littoralis
Olearia ilicifolia
Cordyline indivisa

56 Transactions – BOTANY VOL. 3
LIST 3. GENERAL. FOREST SPECIES (Continued)
A. Upper Limit 2,000-3,000ft B. Upper Limit 3,000-4,000ft
UNDERSHRUBS
Neomyrtus pedunculata
Neopanax anomalum
Gaultheria antipoda
Cyathodes fasciculata
C. juniperina
Alseuosmia pusilla
Coprosma microcarpa
C. robusta
C. banksii
C. rhamnoides
C. lucida
Brachyglottis repanda
Pseudowintera colorata
Aristotelia fruticosa var.
Myrsine divaricata
Coprosma parviflora
C. foetidissima
C. tenuifolia
Hebe corriganii
FLOOR
Todea hymenophylloides
Gleichenia cunninghamii
Hymenophyllum multifidum
Asplenium bulbiferum
Blechnum capense
B. discolour
B. vulcanicum
B. Fluviatile
Astelia solandri
A. nervosa var. sylvestris
Microlaena avenacea
Todea superba
Dicksonia lanata
Paesia scaberula
Enrage parviflora
Libertia pulchella
Chionochloa conspicua
Blechnum minus
EPIPHYTES
Hymenophyllum dilatatum
H. rarum
H. demissum
Phymatodes diversifolium
Grammitis heterophylla
Asplenium flaccidum
Metrosideros diffusa
Collospermum hastatum
Hymenophyllum malingii (confined to Libo-cedrus)
H. sanguinolentum
Elytranthe tetrapetala (Parasite)
LIST 4. CENTRAL. RIVER BANK
MAT PLANTS
Helichrysum bellidiodes 4
Raoulia tenuicaulis var. dimporpha 4
R. hookeri var. albosericea 3
Pratia angulate 3
Muehlenbeckia axillaris 3
Raoulia glabra 2
Epilobium melanocaulon 2
E. microphyllum 1
Myosotis pygmaea var. drucei 1
Muehlenbeckia ephedroides 1
SHRUBS
Hebe stricta var. stricta 3
Coriaria arborea 3
Helichrysum glomeratum 3
Cortaderia sp. 3
Leptospermum scoparium loc. 2
L. ericoides loc. 2
Coprosma rugosa loc. 3
TREES
Aristotelia serrata 4
Fuchsia excorticate 4
Hoheria sexstylosa 3
Nothofagus solandri var. solandri 3
Sophora tetraptera 3
RIVER CLIFF
Gnaphalium keriense 4
Ourisia macrophylla var. robusta 3
Parahebe catarractae 3
Blechnum patersonii 3
B. vulcanicum 2
Phormium colensoi 3
Senecia latifolius 3

No. 3 ELDER – Vegetation of the Ruahine Range 57
LIST 5A. CENTRAL. HERBS OCCURRING IN TUSSOCK
Recorded above 5,500ft
Chionochloa pallens (snowgrass)
Celmisia spectabilis
C. incana
Coprisma pumila
Gentian bellidifolia
Anosotome aromatic
Oreomyrrhis colensoi
Lycopodium fastigiatum
Recorded above 5,500ft
Astelia cockaynei
Aciphylla colensoi
Coprosma cheesemanii
Ranunculus insignis
R. clivalis
Chrysobactron hookeri
Epilobium chloraefolium
Oxalis lacteal
Recorded only below 5,000ft
Chionochloa rubra (red tussock)
Ourisia colensoi
Taraxacum magellanicum
Epilobium pernitens
Polystichum vestitum
Viola cunninghamii
V. lyallii
Wahlenbergia pygmaea
Coprosma depressa
Phormium colensoi
Myrsine nummularia
Aporostylis bifolia
Blechnum penna-marina
Forstera bidwillii
Plantago lanigera
Aciphylla glaucescens
Celmisia gracilenta
LIST 5B. CENTRAL. SHRUBS OCCURRING IN TUSSOCK
Recorded above 5,500ft
Hebe subsimilis
H. odora
Recorded above 5,000ft
Senecio bidwillii
Podocarpus nivalis
Hebe venustula
Dracophyllum recurvum
Recorded only below 5,000ft
Dacrydium bidwillii
Cassinia vauvilliersii
Olearia colensoi
LIST 5C. CENTRAL. FELL FIELD
Recorded above 5,500ft
Phyllachne colensoi
Leucogenes leontopodium
Raoulia grandiflora
Senuio lagopus
Euphrasia revolute
Recorded above 5,000ft
Parahebe spathulata
P. hookeriana var. olsenii
Ourisia caespitosa
Penta chondra pumila
Myosotis pygmaea
Euphrasia zelandica
Helichrysum bellidioides
Geum parviflorum
Recorded only below 5,000ft
Gaultheria depressa
G. colensoi
Dacrydium laxifolium
Euphrasia cuneata
Wahlenbergia pygmaea
LIST 6. CENTRAL. SCREE
A. On rubble at head of mature scree
Ranunculus insignis
Claytonia australasica
Myosotis pygmaea
Ourisia caespitose
Parahebe hookeriana var. olsenii
B. On running shingle
Epilobium pychnostachyum
E. glabellum
Schizeilema colensoi
Parahebe spathulata (rare)
Senecia glaucophyllus var. raoulii (rare)
S. glaucophyllus var. discoideus (rare)
C. At foot of stabilizing scree
Hebe truncatula

58 Transactions – BOTANY VOL. 3
LIST 7. CENTRAL. SUBALPINE SCRUB below 5,000 ft
(Above 5,000ft cf. List 5B)
Olearia colensoi
Pimelea buxifolia
Gaultheria subcorymbosa
Hebe truncatula
Neopanax colensoi
Pittosporum rigidum
Senecio eleagnifolius
Myrsine divaricata
Dracophyllum filifolium
Phyllocladus alpinus
Coprosma parviflora
C. pseudocuneata
C. foetidissima
LIST 8. NORTHERN. NE PLATEAU SHRUB UNDERSTOREY
Coprosma parviflora 4
Pittosporum rigidum 4
Chionochloa conspicua 4
Coprosma pseudocuneata 3
Dracophyllum filifolium loc. 3
Myrsine divaricate 2
Neopanax simplex 2
Phyllocladus alpinus 2
Hebe corriganii loc. 2
Olearia colensoi loc. 2
Griselinia littoralis 1
Senecio elaeagnifolius (adult) 1
Senecio elaeagnifolius (seedlings) 3
LIST 9. BOG PLANTS
A. General Ruahine Distribution
Sphagnum spp.
Carpha alpine
Schoenus pauciflorus
Except NE Ruahine: Geum leiospermum
Except NW Ruahine: Astelia linearis var. novae-zelandiae
Oreobolus pectinatus
Hypolaena lateriflora
Drosera arcturi
Dacrydium bidwillii
B. Northern and Western Bogs
Chionochloa rubra (red tussock)
Except Whanahuia: Cyathodes pumila
Liparophyllum gunnii
Potamogeton suboblongus
Drosera spathulata
Herpolirion novae-zelandiae
Oreostylidium subulatum
C. Northern Bogs only
Gleichenia circinata
Abrotanella caespitose
Drosera pygmaea
D. North-western Bogs only
Lycopodium ramulosum
Ranunculus ternatifolius
R. depressus var. stewartiae
Drapetes laxus
Nertera balfouriana
Hypsela rivalis
Myosotis ternericaulis
Euphrasia disperma
Utricularia novae-zealandiae
Myriophyllum pedunculatum (and in Tupari Bog)
LIST 10. NORTHERN. RIVER GORGE
Senecia bansii
Pittosporum ralphii
Taucridium parvifolium
Angelica rosaefolia
Chielanthes sieberi
C. distans
LIST 11. NORTHERN. INDUCED SCRUB
A. Manuka scrub (below 3,500ft)
Leptospermum scoparium 5
L. ericoides 4
Olearia furfuracea 4
Phormium colensoi 4
Cassinia vauvilliersii 4
Dracophyllum filifolium 4
Pimelea longifolia 2

No. 3 ELDER – Vegetation of the Ruahine Range 59
B. Subalpine scrub
Dracophyllum subulatum 4
Hymenanthera crassifolia 3
Epacris alpine 3
Olearia furfuracea 3
Pimelea aridula 2
Hebe tetragona 2
Griselinia littoralis 2
Leptospermum scoparium 2
LIST 12. NORTHERN. LIMESTONE CLIFF
Phormium colensoi
Asplenium anomodum
Cystopteris fragilis
Hymenanthera crassifolia
Pimelea aridula
Myosotis petiolate var. petiolate
M. eximia
Parahebe catarractae (broad-leaved form)
LIST 13. WESTERN. AORANGI PODOCARP FOREST
Canopy
Podocarpus totara 4
P. spicatus 4
Gymnelaea (Olea) cunninghamii 4
Dacrydium cupressinum 1
Sub-canopy
Aristotelia serrata loc. 4
Coprosma linariifolia 2
Griselinia littoralis 1
Melicytus ramiflorus 2
Paratrophis microphylla 2
Lophomyrtus obcordata 2
Lianes
Clematis foetida
Rubus schmidelioides
Rubus cissoids
R. squarrosus
Epiphytes
Phymatodes diversifolium
Astelia solandri (A. Cunn.)
Shrubs
Brachyglottis repanda loc.4
Coprosma rhamnoides 3
C. crassifolia 3
C. virescens 1
C. rubra
Melicytus micranthus
Taucridium parvifolium
Corokia cotoneaster
Urtica ferix
Floor
Microlaena polynoda
Urtica incisa
Parietaria debilis
LIST 14. SOUTHERN. RIMU-RATA FOREST
Below 1,200ft
Laurelia novae-zelandiae
Micropiper excelsum
Lophomyrtus bullata
Sophora sp.
Alectryon excelsum
Pseudopanax edgerleyi
Geniostoma ligustrifolium
Myoporum laetum
Rhopalostylis sapida
Below 2,000ft
Cyathea dealbata
C. medullaris
Beilschmiedia tawa
Hedycarya arborea
Knightia excels
Elaeocarpus dentatus
Myrsine australis (suckering from roots)
Coprosma tenuifolia
C. australis
C. lucida
Solanum aviculare

60 Transactions – BOTANY VOL. 3
Lianes
Blechnum filiforme
Clematis paniculata
Metrosideros fulgens
M. perforate
Metrosideros diffusa
M. Colensoi var. pendens
Parsonia capsularis
Ripogonum scandens
Epiphytes
Pittosporum cornifolium
Senecia kirkii
Dendrobium cunninghamii
Earina mucronata
Earina autumnalis
Astelia solandri (A. Cunn.)
Collospermum microspermum
C. hastatum
Up to 2,300ft
Podocarpus spicatus
P. totara
Dacrydium cupressinum
Metrosideros robusta
Gymnelaea cunninghamii
G. lanceolate
G. montana
Undershrubs
Pseudowintera axillaris
Melicytus ramiflorus
Schefflera digitate
Brachyglottis repanda
To and beyond 2,300ft
Podocarpus ferrugineus
Fuchsia excorticate
Aristotelia serrata
Olearia rani
LIST 15. SOUTHERN. KAMAHI FOREST
Weinmannia racemosa 5
Podocarpus hallii 2
Libocedrus bidwillii loc.2
Dacrydium biforme loc.3
Undershrubs
Pseudowintera colorata 4
Cyathea smithii 3
Griselinia littoralis 2
Myrsine salicina 2
Neomyrtus pedunculata 2
Coprosma foetidissima 2
Floor
Polystichum vestitum 2
Blechnum discolor 3
Histiopteris incisa 4
Uncinia spp. 3
Libertia pulchella 23
Enrage parviflora 2
Todea superba 2
Blechnum nigrum loc.1
LIST 16. SOUTHERN. OPOSSUM BROWSE
Near Extermination
Metrosideros robusta
Weinmannia racemose
Fuchsia excorticate
Alectryon excelsum
Dicksonia squarrosa
Metrosideros fulgens
Neopanax arboretum
N. colensoi
Pseudopanax edgerleyi
Hard Browsed
Elaeocarpus dentatus
Aristotelia racemosa
Hoheria sexstylosa
Myrsine salicina
Senecia elaegnifolius
Neopanax simplex
N. simplex var. sinclairii
Podocarpus hallii
Browsed but Resistant
Brachyglottis repanda
Beilschmiedia tawa
Ripogonum scandens
Melicytus ramiflorus
Schefflera digitate
Pseudowintera colorata

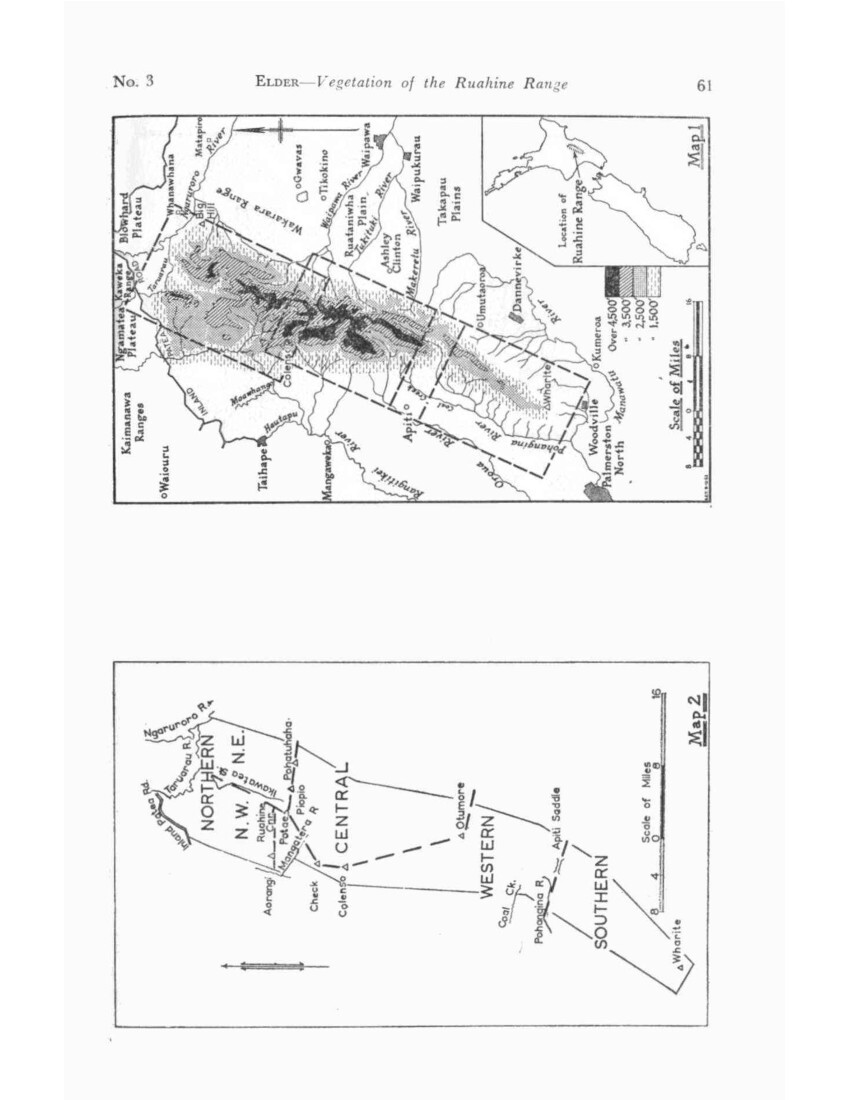
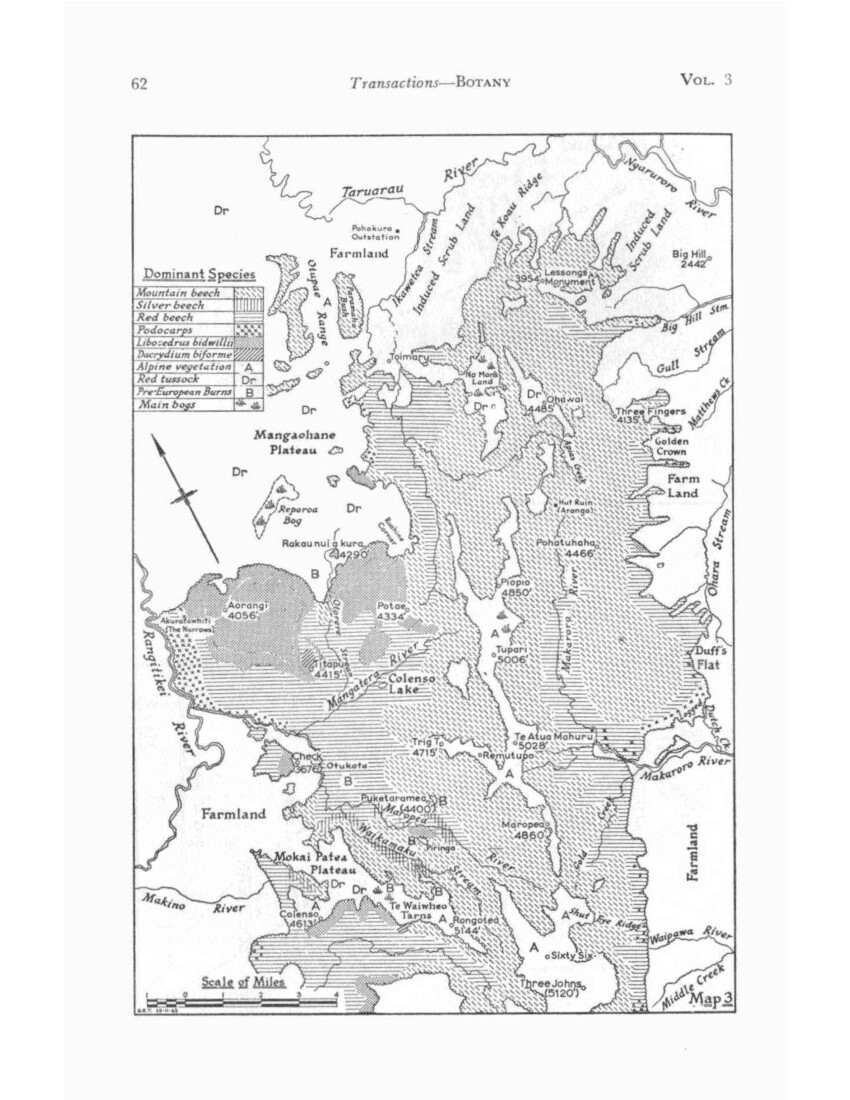
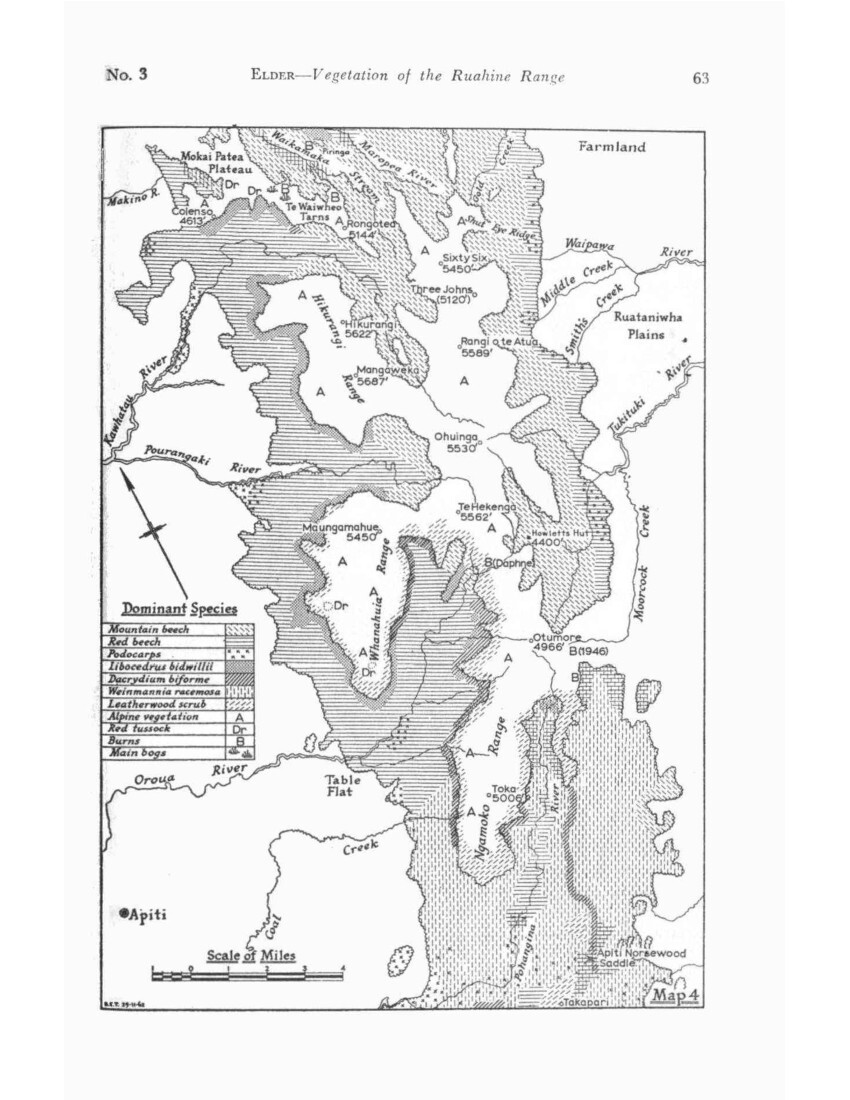
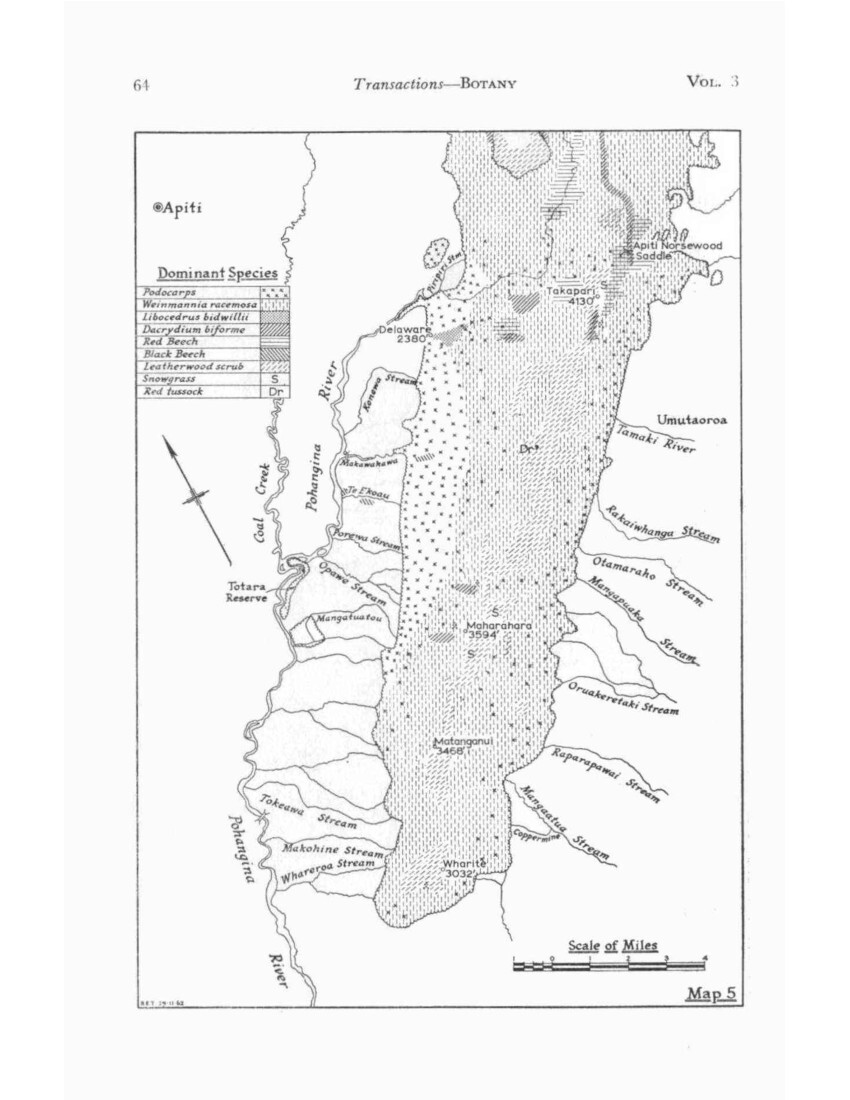
No. 3 ELDER – Vegetation of the Ruahine Range 65
REFERENCES
ALLAN, H. H., 1961. Flora of New Zealand, Vol. 1. Govt. Printer, Wellington
ASTON, B. C., 1914. Flora of Ruahine Mountain Chain. Trans. N.Z. Inst. 46:40.
BAGNALL, A. G., and PEDERSEN, G. C., 1948. William Colenso. Reed, Wellington.
BILLINGS, W. D., and MARK, A. F., 1957. Factors Involved in Persistence of Montane Treeless Balds. Ecology 38(1):140-142.
CHALLIES, C. N. Notes on Birds of the Ruahine Ranges (in preparation).
COLENSO, W., 1884. In Memoriam. Napier.
CUNNINGHAM, A., 1962. Notes on Carbon Fragments in Taupo Ash. N.Z. J. of Botany 2(2), 107-119.
DRUCE, A. P., 1957. Vegetation of Mt. Kaiparoro. Wgtn. Bot. Soc. Bull. 29: 7-13.
ELDER, N. L., 1959. Vegetation of the Kaweka Range. Trans. Roy. Soc. N.Z. 87: 9-16.
ELDER, N. L., 1962. Vegetation of the Kaimanawa Ranges. Trans. Roy. Soc. N.Z. Botany 2(1): 1-37.
ELDER, N. L., 1963. Evidence of Climatic Change in the North Island. N.Z. Ecol. Soc. Proceedings, 10, 45-48.
ELLIS, L. J., 1960. 1959 – 60 Makaroro Assessment. N.Z. Forest Service Report (Unpublished.)
GRANT-TAYLOR, T. L., and RAFTER, T. A., 1962. N.Z. Radiocarbon Age Measurements No. 5. N.Z. J. of Geol. 5(2): 388-390.
GREENWOOD, R. M., 1949. Totara Reserve. Wgtn. Bot. Soc. Bull. 21:2-7
GUTHRIE-SMITH, 1953. Tutira (3rd ed.) Blackwood, Edinburgh, 336-8.
HOLLOWAY, J. T., 1954. Forests and Climates in the South Island of New Zealand. Trans. Roy. Soc. N.Z. 82 (2): 329-410.
KEAN, R., and NEWCOMBE, F. L., 1937. Investigation of Portion of Ruahine Range. Dept. Internal Affairs Ann. Report., 1937.
KIDDIE, D. G., 1962. The Sika Deer (Cervus Nippon) in New Zealand. N.Z. Forest Service, Information Series No. 44.
Geology of Wakarara Range. N.Z. J. of Geol. 1(1), 76 – 91.
tonic History of N.Z. N.Z. J. of Geol 2(1): 1 – 55.
MARK, A. F., 1958. Ecology of S. Appalachian Grass Balds. Ecol. Monographs. 28:293-366.
MASTERS, L., 1960. Back Country Tales. Hart, Hastings.
MOAR, N. T., 1956. Peat on Mokai Patea, Ruahine Range. N.Z. J. Sci. & Tech. 37 (A) (5): 419-426.
– 1961. Pollen Diagrams from W. Ruahine Range. N.Z. J. Sci. 4 (2): 350-359.
MOLLOY, B. P. J., et al., 1963. Distribution of Subfossil Forest Remains, Eastern South Island. N.Z. J. of Botany, 1(1): 68-77.
MORRIN, T. 1956. Mangaohane Station. Hastings Hist. Sect. Records.
PHILLIPPS, W. J., 1959. Last (?) Occurrence of Notornis in North Island. Notornis. 8(4): 93-94.
PRACEY, L., 1948. Internal Affairs reports. (Unpublished.)
– 1949. Internal Affairs reports. (Unpublished.)
ROBBINS, R. G., 1962. The Podocarp-Broadleaf Forests of New Zealand. Trans. Roy. Soc. N.Z. (Botany) (1)5: 33-75.

66 Transactions – BOTANY VOL. 3
THODSEN, -, 1957. Early Settlement of Norsewood. Hastings Hist. Sect. Records.
WARDLE, P., 1960. The Subalpine Scrub of the Hokitika Catchment, Westland. Trans. Roy. Soc. N.Z. 88: 47-61.
– 1962. Subalpine Forest and Scrub, Tararua Range. Trans. Roy. Soc. N.Z. Bot. 1(6): 77-89.
– 1963a. Evolution of New Zealand Flora. N.Z. J. of Botany 1(1) 3-17
– 1963b. Growth Habits of Subalpine Shrubs and Trees. N.Z. J. of Botany 1(1) 18-47
WIDDOWSON, D. 1960. Beech Regeneration and Deer in the N.E. Ruahine Range. N.Z. J. Forestry, 8: 335-336.
WILLETT, R. W. 1950. N.Z. Pleistocene Snowline. N.Z. J. Sci. & Tech. 32(1): 18-48.
WODZICKI, K. A., 1950. Introduced Mammals of N.Z. Dept. Sci. & Indust. Research Bull. 98.
ZOTOV, V. D., et al., 1938. Outline of Vegetation and Flora of Tararua Mountains. Trans. Roy. Soc. N.Z. 68: 259-324.
– 1963. Synopsis of the Grass Subfamily Arundinoideae in N.Z. N.Z. J. of Botany 1(1), 78-136.
N.L. ELDER,
61 McHardy Street,
Havelock North.

{kind=link}
{kind=link}
Original digital file
CollinsGW1994-16_Vegetation_Ruahine.pdf
Non-commercial use

This work is licensed under a Attribution-NonCommercial 3.0 New Zealand (CC BY-NC 3.0 NZ).
Commercial Use
Please contact us for information about using this material commercially.Can you help?
The Hawke's Bay Knowledge Bank relies on donations to make this material available. Please consider making a donation towards preserving our local history.
Visit our donations page for more information.
Description
Surnames in this publication –
Allan, Aston, Bagnall, Batley, Beamish, Billings, Campbell, Challies, Colenso, Cookson, Cunningham, Don, Druce, Ellis, Forst, Grant-Taylor, Greenwood, Hamlin, Holloway, Hutchinson, Johnson, Kean, Kiddie, Kingma, Klein, Linn, Logan, Mackenzie, Mark, Masters, Mill, Moar, Molloy, Morrin, Newcombe, Oerst, Oliver, Pedersen, Phillipps, Poole, Port, Pracey, Rafter, Rich, Richardson, Robbins, Romanes, Severinsen, Sixtus, Soland, Thodsen, Wardle, Widdowson, Willett, Wodzicki, Zotov
Subjects
Tags
Business / Organisation
Forest Research InstituteFormat of the original
Book paperbackDate published
1965Accession number
636625Supporters and sponsors
We sincerely thank the following businesses and organisations for their support.




Do you know something about this record?
Please note we cannot verify the accuracy of any information posted by the community.